Leanne Zrum and Brenda J. Hann
Department of Zoology, University of Manitoba
Winnipeg, Manitoba, Canada R3T 2N2
| The impact of invertebrate and
vertebrate predation on littoral zooplankton in a wetland ecosystem Leanne Zrum and Brenda J. Hann Department of Zoology, University of Manitoba Winnipeg, Manitoba, Canada R3T 2N2 |
|
The importance of predation by planktivorous fish has been demonstrated to be influential in the top-down control of zooplankton communities in both the pelagic and littoral zones of freshwater ponds and lakes (Doolittle 1982; Pont et al. 1991; Diehl 1992). Within the littoral zone, submersed vegetation affords zooplankton protection from fish predation by providing a refugium (Straskraba 1965; Crowder and Cooper 1982). Although fish predation may become more limited within the littoral zone, the littoral zooplankton will be impacted also by invertebrate predation pressure (Goulden 1971; Daggett and Davis 1974). Thus, when investigating top-down control of a zooplankton community in a marsh system which consists largely of littoral zone, different sources of predation pressure must be considered.
The littoral zooplankton in freshwater wetlands, such as Delta Marsh, are exposed to predation by invertebrate and vertebrate predators. Among invertebrate predators are the aquatic representatives of different invertebrate groups, primarily larval insects, for example, dragonfly naiads, damselfly naiads, dytiscid larvae, notonectid nymphs, corixid nymphs, ephemeropteran larvae, as well as Mesostoma sp., Chaetogaster sp., Hydra sp., and water mites. Vertebrate predators include salamanders, e.g., Ambystoma tigrinum, planktivorous fish, e.g., fathead minnow, spottail shiner, and five- and ninespine stickleback, piscivorous fish, e.g., northern pike and yellow perch, and detritivorous/omnivorous fish, e.g., carp and black bullhead (Schneider 1983).
Planktivorous fish are primarily visual predators which tend to feed in a size-selective fashion, i.e. they selectively remove large individual zooplankton when foraging, allowing the small zooplankton which escape predation to become the dominants (Brooks and Dodson 1965; Hall et al. 1976). Invertebrate predators include both tactile and visual predators, with very different feeding strategies in comparison to fish. Predation by invertebrates has been shown to be sufficient to eliminate smaller zooplankton species because they are easier to capture and ingest, thus being size-limited or size-selective in a direction opposite to that achieved by fish predation (Dodson 1974; Williamson 1987).
The purpose of this study was to investigate the effects of differential predation by planktivorous fish species and invertebrates on the species richness, composition, diversity, and size structure of the littoral zooplankton community (Cladocera, Copepoda, Rotifera) over the open-water season in two contrasting areas of the Delta Marsh. In Crescent Pond (CP), a fishless location, the zooplankton community is exposed to invertebrate predation, whereas in Blind Channel (BC), with numerous fish species, both vertebrate and invertebrate predators are abundant. We considered the hypothesis that top-down control via size-selective predation is of primary importance for determining the food-web constituents in these two distinct study areas.
Crescent Pond is a small, self-contained pond with no direct connection to Lake Manitoba. Blind Channel, however, is a long, meandering channel with indirect exchange of water with the lake via Cram Creek. The position of Crescent Pond in Delta Marsh offers it some protection from the effects of wind. The more exposed position of Blind Channel results in a greater amount of wind-induced turbidity. An additional source of turbidity in Blind Channel is the activity of the detrivorous/omnivorous fish which feed using their snouts to disturb the top layer of sediment, resuspending it in the process. The lack of turbidity in Crescent Pond provides the submersed vegetation with an increased amount of irradiance, permitting the earlier establishment of aquatic vegetation in Crescent Pond in comparison to Blind Channel. Thus, the water clarity of Crescent Pond is much greater than that for Blind Channel throughout the summer months, resulting in two distinct aquatic habitats.
Three transects were established along the north margin of Crescent Pond and in Blind Channel near the entrance to Canoe Ditch. Each transect consisted of a nearshore site, located at the edge of the Typha sp. in <<1 m of water, and an offshore site, located in approximately 1 m of water. Sites along each transect were 5 m apart and transects were 20 m apart. The water depth fluctuated (± 5 cm) throughout the summer in response to varying weather conditions.
Sampling of BC and CP was carried out on a weekly basis for 14 consecutive weeks from 24 May to 24 August 1994. The water column was sampled for zooplankton using a transparent acrylic cylinder 50 cm in length and 5.5 cm in diameter. A 4 L volume was then filtered through a conical net with a mesh size of 80 µm. Samples were preserved with formalin and the volume of each was standardized to 20 mL. The water column samples for week 14 for BC were collected through dense macrophyte growth resulting in anomalously high densities of organisms. As this did not accurately represent the true densities in the water column, these samples were not included in the analyses.
Zooplankton were identified to species using various standard references including Pennak (1978), Edmondson (1959), and Smith and Fernando (1978). The Shannon diversity index (H’) was used to determine the species diversity for cladocerans found in CP and BC water column samples over the 14-week sampling period.
Most aquatic predators are size-selective so body size reflects prey availability and food consumption potential in predator-prey relationships. Thus, analysis of the size structure of the zooplankton community was carried out. After each water column sample was counted, the total number of organisms in each was determined, and the proportion of each species contributing to 100% was calculated. On the basis of these proportions, length measurements for 100 individuals were obtained using the digitizing program ZOOBENTH. Length of cladocerans was determined from the top of the head to the base of the shell spine (if present) or postero-dorsal angle of the carapace. Length of copepods was determined from the top of the cephalothorax to the base of the caudal rami. Measurements for identified cladoceran and copepod species were taken from weeks 1, 4, 8, 11 and 14 for BC and CP water column samples. The 100 measurements per site per week were pooled to produce a total of 600 lengths from which the length-frequency histograms for the two locations were assembled. By standardizing the size intervals used to construct the histograms we may observe the size-frequency distribution for micro-invertebrates for either location at a particular point in time, observe changes in the size-frequency distribution for either location over time, and observe differences between the two locations at a particular point in time and over time.
A total of 20 species of cladocerans was found in CP, but only 18 species were found in BC during the course of the study. In addition, four species of cyclopoid copepod and one species of calanoid copepod were found in both locations throughout the sampling period. One harpacticoid copepod was reported from BC. Rotifers were counted but not identified. The temporal distribution of species and species richness per week is provided for CP and BC in Tables 1 and 2, respectively.
| Table 1. The temporal distribution of species and species richness per week for Crescent Pond water column, 1994; results for sites 1 through 6 pooled per week. | ||||||||||||||
Week |
||||||||||||||
| Taxon | 1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
| CLADOCERA | ||||||||||||||
| Alona sp. | P |
P |
P |
P |
P |
|||||||||
| Alonella sp. | P |
P |
P |
P |
P |
|||||||||
| Bosmina longirostris | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|
| Camptocercus sp. | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
||||
| Ceriodaphnia sp. | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| Chydorus sp.1 (sm.) | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| Chydorus sp. 2 (lg.) | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|
| Daphnia magna | P |
|||||||||||||
| Daphnia pulex | P |
P |
P |
|||||||||||
| Daphnia rosea | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|||
| Diaphanosoma sp. | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| Eurycercus sp. | P |
P |
P |
|||||||||||
| Graptolebris sp. | P |
P |
P |
P |
||||||||||
| Kurzia latissima | P |
|||||||||||||
| Leydigia leydigi | P |
|||||||||||||
| Pleuroxus denticulatus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
||
| Pleuroxus procurvus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
||||
| Polyphemus pediculus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
||
| Scapholebris sp. | P |
P |
P |
P |
P |
|||||||||
| Simocephalus vetulus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|
| COPEPODA | ||||||||||||||
| Cyclopoida | ||||||||||||||
| Cyclops bicuspidatus thomasi | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| M/F | M |
M/F |
M |
M/F |
F |
M |
F |
M |
M/F |
M/F |
F |
F |
||
| Cyclops varicans rubellus | P |
P |
P |
P |
P |
P |
P |
P |
||||||
| M/F | M/F |
M/F |
M |
M |
M/F |
F |
M |
M/F |
||||||
| Eucyclops agilis | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|||
| M/F | M/F |
M/F |
M/F |
M/F |
F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
|||
| Macrocyclops albidus | P |
P |
P |
P |
P |
P |
P |
P |
P |
|||||
| M/F | F |
F |
F |
M |
M/F |
M |
M/F |
M/F |
||||||
| Calanoida | ||||||||||||||
| Diaptomus nudus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| M/F | F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
| ROTIFERA | ||||||||||||||
| Asplanchna sp. | P |
P |
P |
P |
P |
P |
P |
|||||||
| TOTAL SPECIES | 8 |
10 |
13 |
17 |
15 |
18 |
16 |
18 |
20 |
21 |
19 |
17 |
19 |
16 |
| Table 2. The temporal distribution of species and species richness per week for Blind Channel water column, 1994; results for sites 1 through 6 pooled per week. | ||||||||||||||
Week |
||||||||||||||
| Taxon | 1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
| CLADOCERA | ||||||||||||||
| Alona sp. | P |
P |
P |
P |
P |
P |
P |
P |
||||||
| Bosmina longirostris | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| Camptocercus sp. | P |
P |
||||||||||||
| Ceriodaphnia sp. | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|
| Chydorus sp.1 (sm.) | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
|
| Chydorus sp. 2 (lg.) | P |
P |
P |
P |
||||||||||
| Daphnia pulex | P |
P |
||||||||||||
| Daphnia rosea | P |
P |
P |
P |
P |
P |
||||||||
| Diaphanosoma sp. | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| Eurycercus sp. | P |
P |
P |
P |
P |
P |
P |
P |
||||||
| Latona sp. | P |
|||||||||||||
| Leptodora kindti | P |
|||||||||||||
| Pleuroxus denticulatus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
||||
| Pleuroxus procurvus | P |
P |
P |
P |
||||||||||
| Polyphemus pediculus | P |
P |
P |
|||||||||||
| Scapholebris sp. | P |
P |
P |
|||||||||||
| Simocephalus serrulatus | P |
P |
P |
P |
P |
P |
P |
P |
||||||
| Simocephalus vetulus | P |
P |
P |
P |
P |
P |
P |
P |
P |
|||||
| COPEPODA | ||||||||||||||
| Cyclopoida | ||||||||||||||
| Cyclops bicuspidatus thomasi | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| M/F | M |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
M/F |
|||
| Cyclops varicans rubellus | P |
P |
||||||||||||
| M/F | M/F |
M/F |
||||||||||||
| Eucyclops agilis | P |
|||||||||||||
| M/F | ||||||||||||||
| Macrocyclops albidus | P |
P |
||||||||||||
| M/F | M |
|||||||||||||
| Calanoida | ||||||||||||||
| Diaptomus nudus | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
| M/F | M/F |
M/F |
F |
F |
M/F |
F |
M |
M/F |
M/F |
M/F |
||||
| Harpacticoida | P |
P |
P |
P |
P |
P |
P |
P |
P |
|||||
| M/F | ||||||||||||||
| ROTIFERA | ||||||||||||||
| Asplanchna sp. | P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
P |
||
| TOTAL SPECIES | 9 |
8 |
12 |
11 |
16 |
15 |
14 |
15 |
16 |
11 |
13 |
9 |
12 |
16 |
Cladoceran species diversity was similar in both sites (Fig. 1) showing low values early in May, then fluctuating around H’ = 1.1 for the remainder of the summer months.
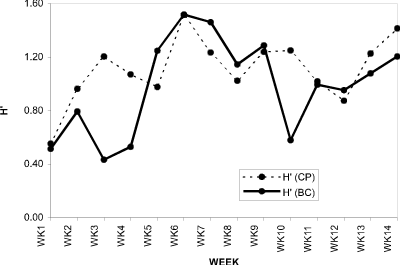
Figure 1. Cladoceran species diversity (H’) for Crescent
Pond and Blind Channel water column, 1994.
Densities for cladocerans, cyclopoid copepods, calanoid copepods and rotifers (Asplanchna) (Fig. 2) are shown for the 1994 season. At week 1 of sampling, the cladocerans were at the lowest density compared to the cyclopoids and the calanoids due to the different strategies employed by these groups for overwintering. At week 5, a large cladoceran peak was observed, with a density an order of magnitude higher than at any other time in the season. The cladoceran density declined rapidly after the bloom and remained consistently less than 100 individuals per liter for the remainder of the sampling period.
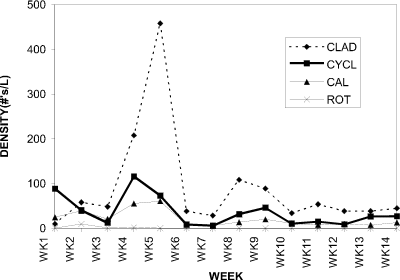
Figure 2. Water column densities of cladocerans (CLAD),
cyclopoid copepods (CYCL), calanoid copepods (CAL), and rotifers
(ROT) for Crescent Pond, 1994.
Both the cyclopoid and calanoid copepods were at a higher initial density in the spring than the cladocerans, but by week 2 cyclopoids had declined to a density comparable to that of the calanoids, both lower in magnitude than the cladocerans. The cyclopoid and calanoid densities remained consistently low (less than 50 individuals per liter) for the remainder of the sampling period.The stability of the copepod numbers is due in part to their exclusively sexual mode of reproduction which limits the rate of their numerical response to fluctuations in food availability.
The densities for individual cladoceran (Fig. 3) and copepod (Fig. 4) species are shown for the 1994 season. Contributing to the very large cladoceran peak at week 5 were Ceriodaphnia dubia, Daphnia rosea and Chydorus spp. Most cladoceran species attained their highest seasonal densities at week 5. A slight recovery was seen at week 8 with C. dubia dominating, but a peak similar to week 5 did not recur. The dominant cyclopoid species was Diacyclops thomasi; Eucyclops agilis took over as the dominant at week 5 only and was equivalent in numbers to D. thomasi at week 6.
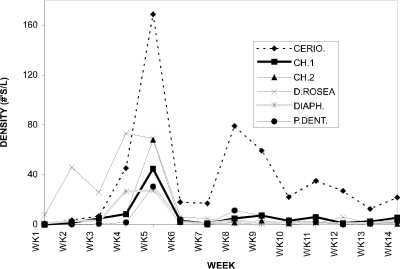
Figure 3. Water column densities of individual cladoceran
species for Crescent Pond, 1994; CERIO. = Ceriodaphnia dubia, CH.1
= Chydorus sp.1, CH.2 = Chydorus sp.2, D.ROSEA = Daphnia
rosea, DIAPH. = Diaphanosoma birgei, P.DENT. = Pleuroxus
denticulatus.
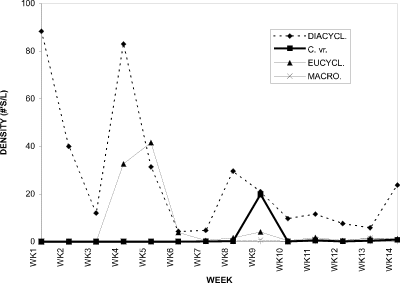
Figure 4. Water column densities of individual cyclopoid copepod
species for Crescent Pond, 1994; DIACYCL. = Diacyclops thomasi,
C. vr. = Cyclops varicans rubellus, EUCYCL. = Eucyclops
agilis, MACRO. = Macrocyclops albidus.
When the relative proportions of the groups were considered, the cladocerans were consistently dominant (Fig. 5). The proportion of cladocerans was initially lower than copepods (calanoids and cyclopoids) at week 1, but increased rapidly to an averaged plateau of approximately 60-70 % for the season. Therefore, although the density of cladocerans (Fig. 2) declined after the peak at week 5, their relative percentage contribution (Fig. 5) remained high until week 13, dropping to 55 %. Copepods did not contribute more than approximately 30 % of the total until week 13 when they constituted approximately 45 % of the total.
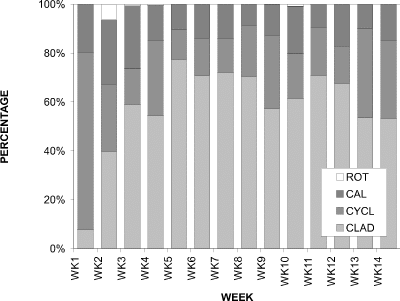
Figure 5. Relative proportions of water column densities of
cladocerans (CLAD), cyclopoid copepods (CYCL), calanoid copepods
(CAL), and rotifers (ROT) for Crescent Pond, 1994.
When the proportions of six individual cladoceran species contributing to the total for the cladoceran community were considered, two dominant patterns emerged (Fig. 6). At week 1 the dominant cladoceran was Daphnia rosea with a proportion of approximately 85 %. As the sampling period progressed, the proportion of Daphnia rosea declined. As this was occurring, Ceriodaphnia dubia was becoming increasingly dominant. By week 7 the proportion of Daphnia rosea had dropped to close to 10 %, while the proportion of Ceriodaphnia dubia had increased to approximately 75%. The contribution by the other four species combined never exceeded 40 %.
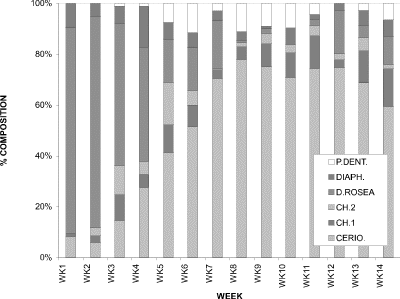
Figure 6. Relative proportions of water column densities of
individual cladoceran species for Crescent Pond, 1994; CERIO. = Ceriodaphnia
dubia, CH.1 = Chydorus sp.1, CH.2 = Chydorus
sp.2, D.ROSEA = Daphnia rosea, DIAPH. = Diaphanosoma
birgei, P.DENT. = Pleuroxus denticulatus.
The cyclopoid copepod assemblage in CP was comprised of four species (Fig. 7). When the relative proportions of each were considered for the sampling period, one species in particular stood out as being dominant, Diacyclops thomasi. For the first three weeks its proportion was 100 %. The only times it contributed less than 63% was at weeks 5, 6, and 9. At week 5 Eucyclops agilis had a proportion of 58 % and was the dominant species. D. thomasi and E. agilis contributed 53 % and 46 %, respectively, with 1 % being attributable to Macrocyclops albidus, at week 6. The species dominating at week 9 included Cyclops varicans rubellus (42 %), along with D. thomasi (48 %); contributing the remaining 10 % are E. agilis (8 %) and M. albidus (2 %). During the intervening weeks of 7 and 8, and 10 through 14, D. thomasi was dominant.
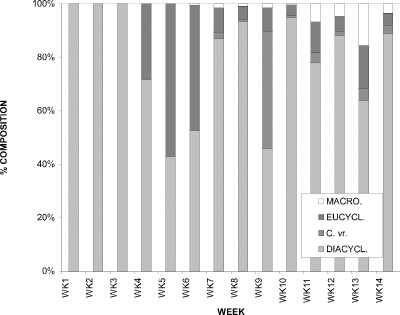
Figure 7. Relative proportions of water column densities of
individual cyclopoid copepod species for Crescent Pond, 1994; DIACYCL.
= Diacyclops thomasi, C. vr. = Cyclops varicans
rubellus, EUCYCL. = Eucyclops agilis, MACRO. = Macrocyclops
albidus.
At week 1 (24 May) (Fig. 8), the histogram peak of 450 µm corresponded to cyclopoid copepods with cladoceran species making a minor contribution. Measurements of calanoid copepods skewed the distribution toward larger-sized organisms. In week 4 (Fig. 9), however, the predominant contributers were now cladoceran species throughout the size range. Cyclopoids again comprised a peak at 450 µm and calanoids at 1200 µm. In addition to calanoids, larger cladocerans (e.g., Simocephalus vetulus) were responsible for the histogram being skewed in the direction of larger-sized organisms. The peak at week 8 (13 July) (Fig. 10) still corresponded to cladoceran species, but their modal size had been reduced to 300 µm. Cyclopoids again formed the modal size at 450 µm and calanoids peaked at 1100 µm. By week 11 (3 August) (Fig. 11), the histogram had become more bimodal with peaks of predominantly smaller cladocerans at 450 µm and calanoid copepods and larger cladoceran species occurring at 1000 µm. A more distinct bimodal distribution was observed at week 14 (24 August) (Fig. 12) with a broad higher frequency peak at 450 µm of cladocerans and cyclopoids and a second lower frequency peak at 1100 µm of calanoids/larger-sized cladocerans. The size-distribution for CP was consistently bimodal, with cyclopoids, Ceriodaphnia dubia, and other cladocerans at a peak of 400-500 µm, and calanoids, and larger cladoceran species (e.g., Daphnia rosea and Simocephalus vetulus) at a peak of 1100 µm.
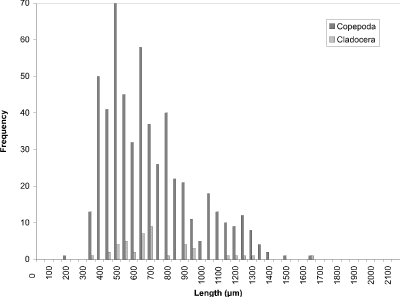
Figure 8. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Cresent Pond water column, May 24 1994
(Week 1).
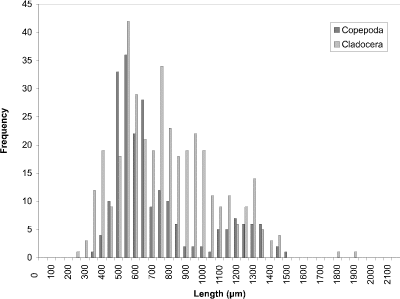
Figure 9. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Crescent Pond water column, June 16
1994 (Week 4).
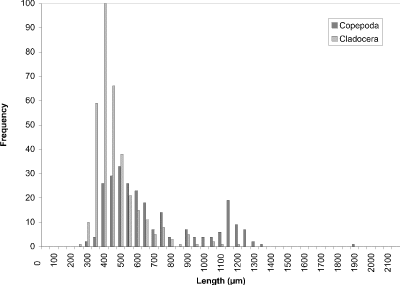
Figure 10. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Crescent Pond water column, July 13
1994 (Week 8).
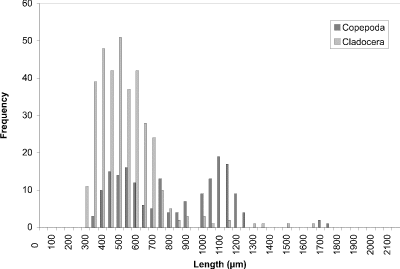
Figure 11. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Crescent Pond water column, August 3
1994 (Week 11).
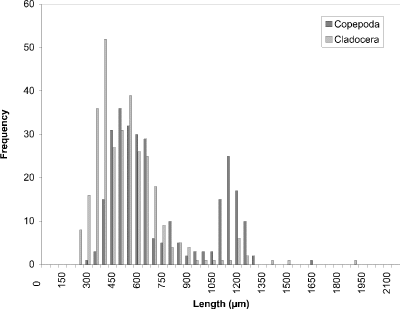
Figure 12. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Crescent Pond water column, August 24
1994 (Week 14).
Figure 13 presents the changes in density (#/L) for the cladoceran, cyclopoid, calanoid and rotifer groups considered in BC. In contrast to CP (Fig. 2), the planktonic rotifer species Asplanchna contributed substantially to the total density of micro-invertebrates counted.
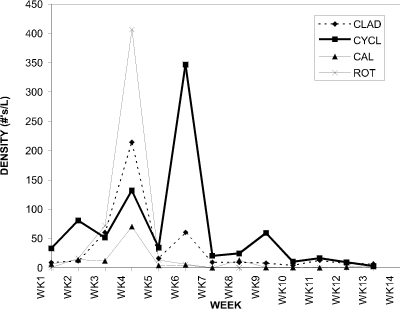
Figure 13. Water column densities of cladocerans (CLAD),
cyclopoid copepods (CYCL), calanoid copepods (CAL), and rotifers
(ROT) for Blind Channel, 1994.
At week 1 the cladocerans were at a very low density, but their numbers increased steadily towards a peak at week 4. A second smaller peak at week 6 was observed before the cladoceran numbers fell and remained at a low density (less than 20 individuals per liter) for the remainder of the sampling period. Aside from the peak at week 4, cladoceran numbers remained below those for the cyclopoid copepods for the entire sampling period. This is the opposite of the situation observed in the cladoceran-dominated CP.
A similar trend to the cladocerans was seen for the rotifers. From a very low density at week 1, the rotifer Asplanchna sp. increased to a very large peak at week 4. Both groups reproduce parthenogenetically/asexually and have short generation times, which may contribute to the similar pattern observed. Since Asplanchna sp. is a predatory rotifer feeding on other rotifers, planktonic crustaceans and colonial algae, it is affected by the availability of different food sources than the cladocerans. However, the reproductive rates of both Asplanchna sp. and the cladocerans are related to the quality and abundance of food, in addition to water temperature.
At week 1 the cyclopoid copepods were the most dense group, with the calanoid copepods being amongst the lowest. The cyclopoid group showed three successively higher peaks at 2-week intervals from week 2 to week 6, then declined to stable densities less than 50 individuals per liter for the rest of the summer. The numbers of calanoid copepods remained low and steady (less than 10 individuals per liter) other than a slight peak at week 4.
The cladoceran peak observed at week 4, with density two orders of magnitude higher than at any other time in the season, consisted overwhelmingly of Bosmina longirostris (Fig. 14). Ceriodaphnia, Daphnia, and Chydorus, species involved in the cladoceran peak at week 5 in CP (Fig. 3), are relatively large when compared to the smaller Bosmina longirostris forming the BC peak at week 4. A second minor peak of Diaphanosoma birgei and Ceriodaphnia dubia was observed at week 6 in BC.
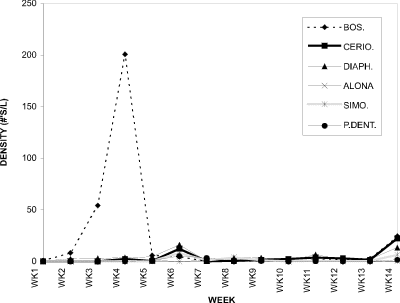
Figure 14. Water column densities of individual cladoceran
species for Blind Channel, 1994; BOS. = Bosmina longirostris, CERIO.
= Ceriodaphnia dubia, DIAPH. = Diaphanosoma birgei,
ALONA = Alona spp., SIMO. = Simocephalus spp., P.DENT.
= Pleuroxus denticulatus.
The dominant cyclopoid in BC was D. thomasi, a carnivorous species (Fig. 15) which preys on other micro-crustaceans, dipteran larvae and oligochaetes (Wetzel 1983). Other cyclopoid copepods and calanoids were present at very low densities. Except for the Asplanchna sp. and Bosmina longirostris peaks at week 4 (Figs. 13, 14), D. thomasi was the dominant microcrustacean in BC for the entire sampling period.
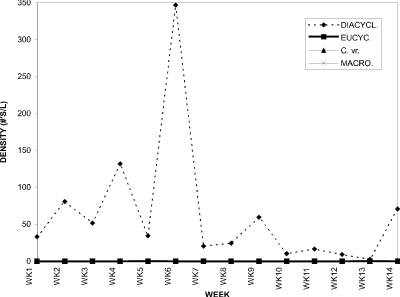
Figure 15. Water column densities of individual cyclopoid copepod
species for Blind Channel, 1994; DIACYCL. = Diacyclops thomasi,
EUCYCL. = Eucyclops agilis, C. vr. = Cyclops varicans
rubellus, MACRO. = Macrocyclops albidus.
In contrast to CP where the cladocerans constituted a major proportion of organisms, in BC they made up no more than approximately 35 % of the community until week 11, when their percent contribution rose to nearly 45 % (Fig. 16). A peak of close to 50 % was seen at week 13, however this higher percent contribution may be misleading as the cladoceran density and densities of all microinvertebrates at week 13 (Fig. 13) were very low. Similarly, the peaks in cladoceran density (Fig.13) at weeks 4 and 6 were not reflected in their percent contribution (Fig.16), as the cladocerans contributed only approximately 35 % at week 4 and 15 % at week 6. This may be due in part to the very large coincident peaks in rotifer (at week 4) and cyclopoid (at week 6) densities (Fig. 13).
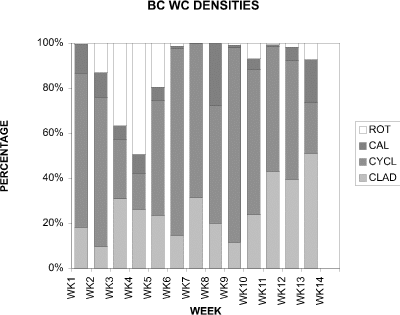
Figure 16. Relative proportions of water column densities of
cladocerans (CLAD), cyclopoid copepods (CYCL), calanoid copepods
(CAL), and rotifers (ROT) for Blind Channel, 1994.
The percent contribution of Asplanchna sp. (Fig.16) followed the same pattern as its density (Fig.13). The large peak in rotifer density in at week 4 corresponded to the large percent contribution by the rotifers on the same date. However, after this peak, the percentage of rotifers declined to a relatively negligible amount for the remainder of the sampling period.
Of the groups considered, the percent contribution of the cyclopoid copepods to the total was the greatest. The percent contribution by cyclopoids at week 1 was comparable in both CP and BC (Figs. 5 and 16); however, this similarity ceased at week 2 when the percentage of cyclopoids in CP declined significantly. Except for weeks 3, 4 and 13, their percentage in BC was close to 60 % or greater of the total. Their peak in density at week 4 (Fig.13) corresponded to their lowest percent contribution of approximately 15 % (Fig.16). This artifact was due in part to the exceptionally high density of the rotifer Asplanchna sp. (Fig.13). The calanoid copepods made a significant contribution to the percentage of organisms in weeks 8 and 13 only, otherwise their percent composition ranged from approximately 2-7 %. When compared to CP, the calanoids of BC fluctuated in percentage to a greater degree; for example, at week 7 (Fig. 16) the calanoids were negligible while the next week their contribution increased to approximately 25%.
Bosmina longirostris increased in proportion from 62 % at week 1 to its peak at week 4 when it comprised 97 % of the six dominant cladoceran species considered in BC (Fig. 17). After week 4 its contribution declined to a low of 3 % at week 7, then gradually increased again to more than 39 % (week 13). Diaphanosoma birgei was consistently present, being dominant at week 6 (35 %), 9 (52 %), and 11 (52 %). Other species dominating at particular weeks were Pleuroxus denticulatus at week 7 (59 %), Simocephalus spp. at week 8 (45 %), and Ceriodaphnia dubia at week 10 (75 %). Therefore, following the early bloom of Bosmina longirostris, the cladoceran community in Blind Channel consisted of a more balanced mix of species, with densities and dominance of individual species fluctuating throughout the season. This was in contrast with the pattern in Crescent Pond (Fig. 6) where the early season cladoceran community was clearly dominated by Daphnia rosea, and the remainder of the season by Ceriodaphnia dubia.
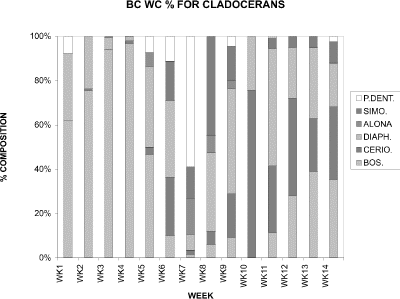
Figure 17. Relative proportions of water column densities of
individual cladoceran species for Blind Channel, 1994; P.DENT. = Pleuroxus
denticulatus, SIMO. = Simocephalus spp., ALONA = Alona spp.,
DIAPH. = Diaphanosoma birgei, CERIO. = Ceriodaphnia
dubia, BOS. = Bosmina longirostris.
D. thomasi comprised 100% of the copepods from week 1 to week 12 (Fig. 18). At week 13, its proportion was 94 %, with the other 6 % being comprised of Eucyclops agilis (4 %) and Macrocyclops albidus (2 %). Cyclops varicans rubellus contributed only 1 % at week 14, with the remainder being D. thomasi.
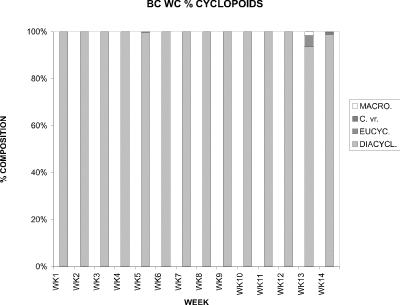
Figure 18. Relative proportions of water column densities of
individual cyclopoid copepod species for Blind Channel, 1994; MACRO.
= Macrocyclops albidus, C. vr. = Cyclops varicans
rubellus, EUCYCL. = Eucyclops agilis, DIACYCL. = Diacyclops
thomasi.
At week 1 (24 May) (Fig. 19), a peak at 450 µm of cyclopoid copepods was observed. Measurements of calanoid copepods skewed the figure toward larger sized organisms. By week 4 (16 June) (Fig. 20) there had been a noticeable shift towards smaller organisms to peak at 250 µm. This peak corresponded to two groups, one comprised of cyclopoids, the second comprised of small cladocerans (e.g., Bosmina longirostris) and the rotifer Asplanchna sp. Calanoids still contributed to the few larger size measurements. A distinctly bimodal histogram was seen at week 8 (13 July) (Fig. 21) with a predominantly cyclopoid peak at 400 µm and a calanoid peak at 1150 µm. The bimodal size-distribution had disappeared by week 11 (3 August) (Fig. 22), being replaced by a single broad peak at 400 µm corresponding to both cladocerans and cyclopoids. The cladoceran portion of the peak was shifted towards smaller sizes, while the cyclopoid portion was shifted towards slightly larger sizes. Calanoids were still the predominant contributers to the larger sized measurements. The distribution observed at week 14 (24 August) (Fig. 23) was the result of slightly offset cladoceran (250 µm) and cyclopoid (450 µm) peaks. Except for week 8, the size distribution in BC was generally unimodal, with substantially fewer large zooplankters represented than observed in CP.
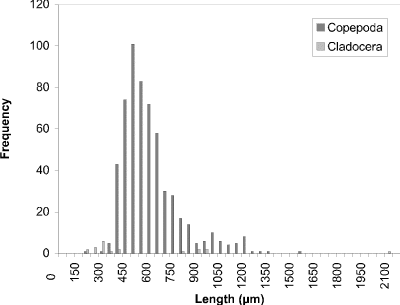
Figure 19. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Blind Channel water column, May 24
1994 (Week 1).
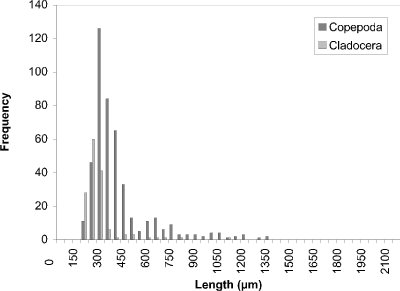
Figure 20. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Blind Channel water column, June 16
1994 (Week 4).
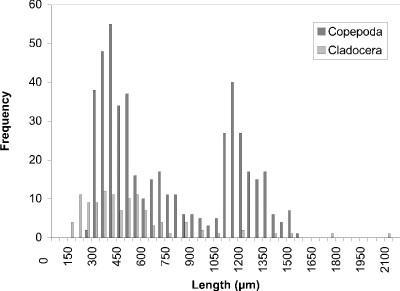
Figure 21. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Blind Channel water column, July 13
1994 (Week 8).
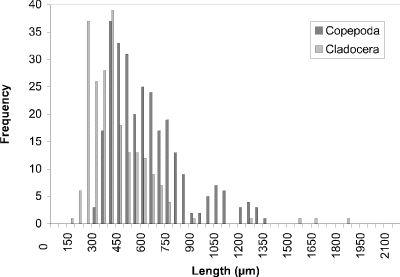
Figure 22. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Blind Channel water column, August 3
1994 (Week 11).
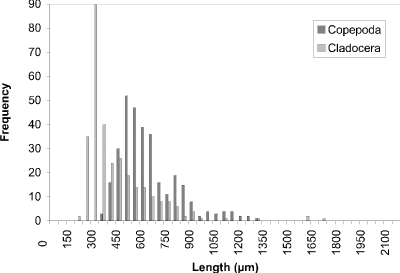
Figure 23. Size-frequency histogram of length measurements of
Copepoda and Cladocera for Blind Channel water column, August 24
1994 (Week 14).
The visual differences between Blind Channel (turbid, sparse macrophytes, and abundant and diverse fish community) and Crescent Pond (transparent, abundant and diverse macrophytes, and no fish) are striking, and consequently one would expect to see very different invertebrate communities in the two study areas. Key differences in zooplankton abundance between the two areas were observed throughout the sampling period. In addition, size structure of the zooplankton community exhibited two distinct patterns as a result of exposure to different types of predation pressure. However, zooplankton community composition in these two distinct habitats of the Delta Marsh became more similar as the summer progressed. The total microcrustacean species pool represented in the two sites in Delta Marsh overlaps considerably with the previously known regional species pool (Smith 1968).
According to size-selective predation theory, larger zooplankton (e.g., Daphnia spp.) should benefit in a waterbody lacking planktivorous fish because these zooplankters are the preferred food of the fish and would be fed on selectively over smaller zooplankton. By removing this particular predation pressure, the abundance of herbivorous zooplankton species present, especially larger species which feed more efficiently than smaller herbivores, would be expected to be higher than in a system with fish present (Hall et al. 1976). In our study, the cladoceran component of the zooplankton community responded most strongly to the differential predation pressure. The abundance of cladocerans (including large Simocephalus and Daphnia) was considerably higher in fishless CP than in BC throughout the season, whereas copepod abundances for the latter half of the season especially were similar at both sites. Cladocerans are typically the preferred prey of planktivorous fish, showing less predator avoidance behaviour than copepods (Drenner et al. 1978).
Despite the markedly higher cladoceran abundances in fishless CP, cladoceran species richness and diversity were similar in the two sites throughout the summer. Cladoceran species composition differed between the two areas, with the occurrence of geographically and numerically rare species being limited to Crescent Pond. The occurrence of these rare cladoceran species in Crescent Pond affected the species diversity index. As species diversity is a combined measure of both species richness and evenness in the distribution of the number of individuals per species, the presence of several sparsely represented species caused the value calculated for H’ to be lower than expected when species richness alone is considered.
Competitive interactions in a speciose zooplankton community may also be involved in the interpretation of the temporal pattern of diversity values in Crescent Pond and Blind Channel if the environmental parameters (physical, chemical, and biotic) that undergo constant temporal variations are considered. Continuously changing environmental parameters (e.g., temperature, turbidity, and food availability) may produce a habitat in which no one invertebrate species maintains a competitive advantage for a sufficient period of time to become the dominant species permanently (Hutchinson 1961; Hall et al. 1970). Environmental conditions in Blind Channel often exhibit rapid temporal changes, resulting in greater habitat variability when compared to Crescent Pond. Consequently, there was temporal instability in the zooplankton community with the dominance of any one cladoceran species in Blind Channel switching frequently, in contrast to the seasonal shift in dominance of cladoceran species in Crescent Pond, with Daphnia rosea dominant for the first half of the summer and Ceriodaphnia dubia dominant for the last half. Lynch (1978) explained a similar Daphnia-Ceriodaphnia species substitution in a fishless pond by seasonal changes in competitive abilities for a shared algal resource.
Hanson and Riggs (1995) conducted a study in a series of wetlands to examine the potential effects of fish predation on wetland invertebrates. They determined that the presence of fathead minnows in particular had an inverse relationship with indices of abundance, biomass and taxon richness of crustaceans and insects. While their observation of elevated abundance (and indirectly biomass) of invertebrates in the absence of planktivorous fish paralleled that found in our study, the pattern for taxon richness was in contrast with our results. However, Hanson and Riggs (1995) did not take into consideration the indirect effect that reduced vertebrate predation on macroinvertebrate predators (insects) has ultimately on the microinvertebrates (crustaceans), or the direct effect invertebrate predation alone has on zooplankton. In addition, the degree of taxonomic resolution used by Hanson and Riggs (1995) is not directly comparable to our study. Their placement of taxa into only two groups (i.e. crustaceans and insects) precludes any interpretation of the response of individual species to varying levels of predation pressure and to differential predation by planktivorous fish and invertebrates.
Densities of cladocerans and copepods in the early spring show different patterns due largely to different evolutionary strategies for adapting to a seasonally variable habitat. Cladocerans overwinter as resting eggs in the sediments, while the copepods tend to overwinter as immature copepodite stages in the water column and/or sediments (Pennak 1978). The strategy of the copepods to overwinter as immature copepodites gives them an initial advantage in the spring over the cladocerans. As a result, there is a time lag in the spring until the cladocerans hatch from their resting eggs during which time the copepods dominate the zooplankton community. In Blind Channel, elevated copepod densities continue throughout the summer with the cyclopoid Diacyclops thomasi being the dominant zooplankter almost every week, except for a brief bloom of the cladoceran Bosmina longirostris. However in Crescent Pond, once the abundance of phytoplankton increases to the extent that sufficient food is available for the asexually reproducing cladoceran populations to expand rapidly, they take over from the copepods as the dominant zooplankters, with Daphnia rosea dominant early in the season and Ceriodaphnia dubia becoming most abundant as the summer progresses.
Although community analyses are informative, analysis of body size, which ignores behavioural, taxonomic, and trophic characteristics, provides very valuable insights into community response to predation. The size-frequency distributions assembled document the shifts in modal sizes for the zooplankton community over the sampling period. The most consistent difference in size distributions is that CP exhibits a bimodal pattern in contrast to the largely unimodal pattern in BC (except in week 8). The second peak of larger organisms is generally absent from BC. At week 1 the measurements obtained for organisms from both areas show a peak at 450 µm, each corresponding to cyclopoid copepods. By week 4 the histograms appear quite different with the peak for Crescent Pond still occurring at 450 µm, but now corresponding to cladocerans and the peak for Blind Channel shifting to a smaller size. This peak at 250 µm in Blind Channel is composed of the small cladoceran Bosmina longirostris and the rotifer Asplanchna sp., in addition to the cyclopoids. It may be inferred that selective fish predation on larger zooplankton has shifted the size distribution to smaller-sized organisms in Blind Channel. The dominance of large cladocerans in Crescent Pond is probably attributable to their superior rapid reproductive capabilities when compared to the copepods, the abundance of algal food and the lack of vertebrate predation pressure. The presence of early instar insect larvae selectively feeding on smaller zooplankton may also be contributing to the maintenance of the peak at a larger size.
The modal size of the cladocerans in Crescent Pond declined slightly at week 8 to 300 µm. The predatory insect larvae may now be larger, feeding on larger prey, and thereby releasing the smaller cladocerans from invertebrate predation. A distinctly bimodal size-distribution is seen in Blind Channel in week 8 only with a cyclopoid peak at 400 µm and a calanoid/larger-sized cladoceran peak at 1150 µm. The occurrence of these two peaks may be the result of the activity of primarily invertebrate predators if planktivorous fish abundance is indeed greatly reduced at this time in the season (Schneider 1983), thereby releasing the large zooplankton from predation. Maximum abundances of planktivores occur typically in May and June in Delta Marsh, then benthivores (e.g., bullheads) migrate into the marsh in August (Schneider 1983).
Submersed macrophytes, at peak biomass in Blind Channel at this time, may also provide a refugium for the large zooplankton (Diehl 1992), while superior swimming skills could allow escape by the cyclopoids from the macroinvertebrate predators. Thus, release from fish predation and increased habitat complexity can result in a bimodality of size-classes of zooplankton, similar to that observed throughout the season in fishless CP.
Predation pressure exerted by fish on zooplankton is typically considered to be more significant than that exerted by invertebrate predators (Diehl 1992) especially in pelagic communities. However, predatory invertebrates may play a more substantial role in zooplankton community dynamics in shallow or littoral ecosystems such as wetlands. Our study has shown that the presence of planktivorous fish does exert a strong degree of predation pressure on the zooplankton community, producing a unimodal size distribution. Strong pressure is also exerted by invertebrate predators, but the higher abundance of zooplankton in a waterbody subject to invertebrate predation alone substantiates that vertebrates are more effective predators than invertebrates (Hall et al. 1970).
This study has focused attention on the roles of both vertebrate and invertebrate predation in zooplankton dynamics in a wetland. Much more now needs to be known regarding seasonal distribution of these predators, and their dietary preferences in order to fine-tune the food web dynamics.
Brooks, J. L. and Dodson, S. I. 1965. Predation, body size, and composition of plankton. Science 150: 28-35.
Crowder, L. B. and Cooper, W. E. 1982. Habitat structural complexity and the interaction between bluegills and their prey. Ecology 63: 1802-1813.
Daggett, R. F. and Davis, C. C. 1974. A seasonal quantitative study of the littoral Cladocera and Copepoda in a bog pond and acid marsh in Newfoundland. Int. Rev. ges. Hydrobiol. 59: 667-683.
Diehl, S. 1992. Fish predation and benthic community structure: the role of omnivory and habitat complexity. Ecology 73: 1646-1661.
Dodson, S. I. 1974. Zooplankton competition and predation: an experimental test of the size-efficiency hypothesis. Ecology 55: 605-613.
Doolittle, W. L. 1982. The nature and cause of the midsummer decline of littoral zooplankton in Lake Itasca, Minnesota. PhD Thesis, University of Tennessee.
Drenner, R. W., Strickler, J. R. and O’Brien, W. J. 1978. Capture probability: the role of zooplankton escape in the selective feeding of planktivorous fish. J. Fish. Res. Bd Can. 35: 1370-1373.
Edmondson, W. T.(ed.) 1959. Freshwater Biology. 2nd Edition. John Wiley and Sons Inc., New York.
Goulden, C. E. 1971. Environmental control of the abundance and distribution of the chydorid Cladocera. Limnol. Oceanogr. 16: 320-331.
Hall, D. J., Cooper, W. E. and Werner, E. E. 1970. An experimental approach to the production dynamics and structure of freshwater animal communities. Limnol. Oceanogr. 15: 839-928.
Hall, D. J., Threlkeld, S. T., Burns, C. W. and Crowley, P. H. 1976. The size-efficiency hypothesis and size structure of zooplankton communities. Ann. Rev. Ecol. Syst. 7: 177-208.
Hanson, M. A. and Riggs, M. R. 1995. Potential effects of fish predation on wetland invertebrates: a comparison of wetlands with and without fathead minnows. Wetlands 15: 167-175.
Hutchinson, G. E. 1961. The paradox of the plankton. Am. Nat. 95: 137-145.
Lynch, M. 1978. Complex interactions between natural coexploiters–Daphnia and Ceriodaphnia. Ecology 59: 552-564.
Pennak, R. W. 1978. Freshwater Invertebrates of the United States. 2nd edition. Wiley and Sons, Inc., New York.
Pont, D., Crivelli, A. J. and Guillot, F. 1991. The impact of three-spined sticklebacks on the zooplankton of a previously fish-free pool. Freshw. Biol. 26: 149-163.
Schneider, F. 1983. Movements of forage fish between Lake Manitoba and Delta Marsh. University of Manitoba Field Station (Delta Marsh) Annual Report 18: 135-145.
Smith, K. and Fernando, C. H. 1978. A Guide to the Freshwater Calanoid and Cyclopoid Copepod Crustacea of Ontario. University of Waterloo Biology Series 18, 76 pp.
Smith, T. G. 1968. Crustacea of the Delta Marsh region, Manitoba. Can. Field-Nat. 82: 120-139.
Straskraba, M. 1965. The effect of fish on the number of invertebrates in ponds and streams. Mitt. Int. Ver. Limnol. 13: 106-127.
Wetzel, R.G. 1983. Limnology. 2nd edition. Saunders College Publ., New York.
Williamson, C. E. 1987. Predator-prey interactions between omnivorous diaptomid copepods and rotifers: the role of prey morphology and behavior. Limnol. Oceanogr. 32: 167-177.