Rhonda L. McDougal and L. Gordon Goldsborough
Department of Botany, University of Manitoba
Winnipeg, Manitoba, Canada
| Responses of wetland algae and macrophytes
to press and pulse additions of inorganic nitrogen and
phosphorus Rhonda L. McDougal and L. Gordon Goldsborough Department of Botany, University of Manitoba Winnipeg, Manitoba, Canada |
|
Wetlands are endangered havens of biodiversity. They are complex and changeable by nature, relying on fluctuating water levels for rejuvenation. Many of the natural cycles, such as fire, flooding and drought, that maintain the health of wetlands are regarded by humans as disastrous events to be controlled. Human activities, including agricultural and road development, urbanization, industrial development and deliberate efforts at preservation and enhancement, often have negative impacts on prairie wetlands (Millar 1989). The degradation and loss of wetlands through human intervention is viewed as one of the major land use issues facing both federal and provincial jurisdictions (Rubec and Rump 1985). Recent estimates place the loss of prairie wetlands in North America at 40% (Canada-United States Steering Committee 1985, cited in Millar 1989). While wetland losses in Canada are not as rapid as in the United States, the pattern of degradation and loss is similar.
Wetlands are defined as shallow-water or littoral-dominated systems (generally < 2 m depth) with ephemeral standing water or continuously waterlogged soil (Goldsborough and Robinson 1996). Globally, wetlands comprise as much as 6% of the total land area (Mitsch and Gosselink 1993). About 14% of Canada’s land area is classified as wetlands, including the extensive northern peatlands (Zoltai 1988). These northern peatlands are particularly important because of their possible role in moderating climate change by providing a major sink for carbon dioxide gas (Kusler et al. 1994). Wetlands provide shoreline buffer zones which limit the damaging effects of waves and floodwaters. Wetlands may also reduce pollution by trapping phosphorus and other chemicals in sediments and providing large populations of aerobic and anaerobic decomposers to break down excess organic matter (Kusler et al. 1994). The Prairie Pothole Region, two-thirds of which occurs in the southern part of the prairie provinces of Canada, is one of the major duck production habitats in North America (Millar 1989). It produces 50% of the duck population in an average year, although it comprises only about 10% of the North American waterfowl breeding area. Wetlands also provide habitat for small mammals, nesting grounds for a wide variety of migratory birds, and spawning grounds for fish, reptiles and invertebrates. Natural shifts in water levels can give rise to the biological diversity of wetland ecosystems. Moisture gradients provide a continuum of growing conditions that can support terrestrial, partially aquatic and fully aquatic vegetation (Kusler et al., 1994). In wetlands where water fluctuation is limited or artificially regulated, other factors such as nutrient loading, grazing pressure and water column stability may interact to affect the structuring of the wetland ecosystem (Goldsborough and Robinson 1996).
Wetzel (1964) suggests that the greatest significance of littoral-dominated ecosystems such as wetlands may be their contribution to primary production for utilization by higher trophic levels. Primary productivity in wetlands is high, ranging from 30 to 80 metric tons per hectare per year (mT/ha/yr) for emergent macrophytes; 5 to 60 mT/ha/yr for epiphytic algae; 2 to 20 mT/ha/yr for submersed macrophytes and 2 to 10 mT/ha/yr for planktonic algae (Wetzel 1983). In particular, the level of the algal standing crop plays an important role in wetland food webs. Because of their small size, algae are more readily consumed than macrophytes by fish and invertebrates (Goldsborough and Robinson 1996). Algae also provide a relatively stable food supply that is available throughout the growing season. The algal standing crop affects the density of invertebrates, which in turn dictates the food supply for waterfowl and other marsh birds (Murkin et al. 1991).
Algal communities in wetlands differ in their ecological requirements and their physical location within the water column. Algal communities considered in this study include phytoplankton, epiphyton, metaphyton and epipelon. Phytoplankton includes algae free-floating in the water column, which may or may not be motile. Epiphyton includes algae growing attached to the surfaces of submersed or emergent vascular and nonvascular plants, termed macrophytes. The term periphyton is used in this study to denote the attached algae growing on the submersed surfaces of experimentally placed acrylic rods, thus differentiating it from the epiphyton for measurement purposes, although its ecological requirements would be similar to those of epiphyton. Metaphyton forms large mats, often composed of filamentous green algae. These mats originate as epiphyton, but detach due to water turbulence and float at or near the water surface due to oxygen trapped within the mats. Epipelon includes algae within the soft sediments which exhibit vertical migration in response to environmental cues such as light.
Goldsborough and Robinson (1996) proposed a model of algal abundance in wetlands that is made up of four alternative stable states dominated alternately by epipelon, epiphyton, metaphyton, or phytoplankton. This model explains the development of a wetland based on the interacting effects of nutrient loading, grazing pressure and fluctuating water levels. According to this model, the four possible stable states that a wetland may attain are (1) the dry marsh state; (2) the open marsh state; (3) the sheltered marsh state; or (4) the lake marsh state. (1) The dry marsh state is characterized by very low water levels that occur following a drought or deliberate drawdown. Because irradiance at the sediment surface is high, epipelon tend to be the predominant algal assemblage. (2) The open marsh state is maintained by periodic natural disturbances in the wetland, leading to epiphyton predominance on the surfaces of submersed and emergent macrophytes. Natural disturbance by benthivorous fish, wind action and high grazing pressure keeps the epiphyton biomass at a level that does not shade the macrophyte substrata to the point of decline. The combined shading effect of the macrophytes and the epiphyton reduces irradiance to the epipelon, keeping epipelon abundance low. Metaphyton is usually knocked back by wind action, and phytoplankton is outcompeted by epiphyton and macrophytes for nutrients in the water column. (3) The sheltered marsh state develops if there is protection from wind action, or there are enough macrophytes in the water column to reduce water movement that causes disaggregation of metaphyton mats. The metaphyton mats that develop shade the macrophytes and the other competing algal communities, thus becoming the dominant assemblage in the sheltered marsh. (4) The lake marsh is characterized by high water levels, abundant nutrients in the water column and low grazing pressure, leading to phytoplankton predominance. Epiphyton, metaphyton and epipelon are not as successful due to low irradiance and low macrophyte abundance. Goldsborough and Robinson (1996) predict that a wetland will proceed in either direction to one of these four stable states, depending on the outcome of the interacting variables of nutrient levels, grazing pressure and water levels. In this study, we looked at one of these interacting variables, the effect of nutrient loading on the structuring of the algal communities.
There has been a great deal of research on the role of increased nutrient supply in the eutrophication of lake ecosystems (Schindler et al. 1971). Increased phosphorus inputs, in particular, have been linked to increased algal growth in lakes (Schindler 1974). Lakes with phosphorus concentrations of 30 to 100 µg/L are defined as eutrophic and above 100 µg/L as hypereutrophic (Wetzel 1983). Under extremely eutrophic conditions, nitrogen utilization may exceed inputs, causing nitrogen to become the growth-limiting nutrient in the system (Wetzel 1983). Most of the current eutrophication research has focused on lakes, which have a significant deep-water or pelagic zone as well as some proportion of littoral zone. The extent to which lake eutrophication research can be generalized to completely littoral-dominated systems such as wetlands is questionable. Nutrient dynamics in wetlands may differ significantly as compared to deeper aquatic ecosystems. Nutrient-rich sediments may be resuspended more frequently as a result of wind action, benthivorous carp, or burrowing invertebrates (Goldsborough 1993). Epipelic algae may produce on oxygen-rich microzone at the sediment-water interface which prevents the efflux of nutrients from the sediments (Goldsborough and Robinson 1985). Waterfowl feces may contribute significantly to the nutrient budget of wetlands, as they do in lakes (Manny et al. 1994). It has been shown that the outcome of resource competition may change along a nutrient gradient and affect the structuring of competing algal communities (Tilman 1977). The extent to which external nutrient loading changes the nutrient gradient and thus affects the structuring of algal communities in wetlands is unknown.
In this study, we perturbed a microcosm of a wetland ecosystem with two patterns of nitrogen (N) and phosphorus (P) additions and measured the growth responses of several algal communities. We hypothesized that the nitrogen and phosphorus additions would cause the wetland ecosystem to progress to one of the alternative stable states described by Goldsborough and Robinson (1996). The periodicity and magnitude of the nutrient additions would alter the composition and relative abundance of epipelon, epiphyton, metaphyton, and phytoplankton. Small regular (press) additions of N and P would encourage either phytoplankton (a “lake marsh” state) or metaphyton (a “sheltered marsh” state) to develop. Phytoplankton may predominate because of its physical position near the top of the water column as first consumer of the regularly added quantities of N and P. Metaphyton may predominate in the sheltered microcosm environment if macrophytes provide sufficient substrata from which to develop. Large spike (pulse) additions of N and P would encourage macrophytes and epiphyton to predominate (an “open marsh” state). The occasional spikes of nutrient would likely not remain in the water column long enough to allow phytoplankton to become dominant. The increased nutrient deposited in the sediments would favor macrophyte growth, thus providing ample substrata for epiphyton abundance.
This study was conducted from 1 May to 31 August 1994. The study site was a stretch of open water in the center of the Blind Channel where the channel was approximately 45 m across. Stands of hybrid cattails, Typha x glauca lined the channel edges and submersed vegetation grew within the channel, including pondweeds (Potamogeton pectinatus, Potamogeton zosteriformis), water-milfoil (Myriophyllum spicatum), hornwort (Ceratophyllum demersum) and stonewort (Chara sp.). The channel was well populated with small fish species, including sticklebacks (Pungitius pungitius), spottail shiners (Notropis hudsonius) and fathead minnows (Pimephales promelas), as well as the larger benthivorous carp (Cyprinus carpio). The channel edges provided habitat for waterfowl and songbirds, as well as beaver and muskrat. The flocculent bottom sediment was a fine-grained black mud that was rich in decomposing organic matter. At the study site, the channel had a relatively flat bottom resulting in uniform water depths of 80 to 100 cm at the beginning of May 1994.
Six large enclosures were installed at the site during the first week of May. Four of the enclosures had been constructed and used for previous research at Delta Marsh (Goldsborough 1991, 1993a). We repaired these four and constructed two new enclosures on the same design. Each enclosure consisted of a 5 m by 5 m wooden frame floating on high-density foam block supports which held the frame just above water level. The plywood top of the frame also provided a 40 cm wide walkway around the enclosure to allow sampling access. A translucent plastic curtain was secured to the inside of each frame and extended through the water column and into the bottom sediments. Metal rebar was enclosed in a pocket at the bottom of the curtain and embedded into the sediments to prevent direct water movement between the enclosure and the surrounding marsh. The total enclosed water volume in each enclosure was about 20,000 L. The six enclosures were bolted together and anchored in the center of the Blind Channel.
We attempted to limit primary consumers, and thereby reduce the number of variables in the experiment, by removing all fish from the enclosures. Minnow traps were immediately placed in each enclosure and emptied on a daily basis. Early in the season, muskrats chewed small holes in each of the curtains, allowing fish to enter the enclosures. The holes were patched immediately and the fish trapped out but not before numerous eggs were laid on the inner side of the curtain. Therefore, the minnow traps were maintained for the duration of the experiment to trap out the fry as they hatched. Several live traps were deployed outside the enclosures to discourage further damage by muskrats.
To provide uniform colonization substrata for periphyton, vertically positioned cylindrical acrylic rods (0.64 cm diameter, 90 cm length) (Goldsborough et al. 1986) were placed in each enclosure on 18 May 1994. The rods were pushed 30 cm into the sediments leaving the uppermost 60 cm of each rod available for algal colonization. One hundred rods were placed in a 10 x 10 grid pattern with equal spacing between each rod and the ones adjacent to it in each enclosure. The rods were positioned far enough from the curtain to avoid abrasion through any contact with it. Prior to placement, we had notched the rods at specific intervals using a bandsaw to provide subsample segments of a predetermined surface area.
Sampling began on 24 May, designated as week 1, and continued on a regular schedule until 26 August (week 14). During weeks 1 to 4, the enclosure site was allowed to recover from the disturbance effect of rod installation. This period also allowed time for initial periphyton colonization of the rods. During this period, sampling was conducted inside and outside the enclosures, as described below, to determine background levels of nutrients and initial measurements of the productivity of algal communities. Nutrient addition began on 20 June, designated as week 5 of the experiment. The manipulated variable was the timing of the nutrient additions. The same total loads of nitrogen and phosphorus were added in two temporal regimes, one designated as the pulse addition and the other designated as the press addition. Two enclosures (labeled C and D) were arbitrarily chosen to receive the pulse nutrient additions, while two more (labeled A and E) were chosen to receive the press nutrient additions. The remaining two enclosures (labeled B and F) were designated as unmanipulated controls. The location of each replicate enclosure was chosen so that no two replicate enclosures were side by side nor on the same side of the Blind Channel. The spacing of the replicates attempted to account for the spatial heterogeneity arising from minor position effects due to prevailing wind direction, water currents and incident angle of light.
The pulse nutrient addition was added at two times over the course of the 15 week experiment, the first one on 20 June 1994 and the second one on 25 July 1994 (Table 1). The press nutrient addition was added every Monday, Wednesday and Friday, beginning 20 June 1994, with a total of 29 additions made over the course of the 15 week experiment. The press addition on the last day of the experiment was not made as no further sampling was carried out after that. At each addition, the measured nutrient chemical for each enclosure was dissolved in 1 L of carbon-filtered water to make an aqueous nutrient solution. At the study site, the aqueous nutrient solution was mixed with 10 L of water from the designated enclosure and sprinkled uniformly from the nozzle of a watering can over the entire surface of the enclosure.
| Table 1. The amount of phosphorus (as NaH2PO4.2H2O) and nitrogen (as NaNO3) added to each of the pulse and press treatment enclosures. The total load reflects the proportion of the inorganic chemical that was elemental N or P. The ratio of total N to total P was 7:1. | |||||
| Designated enclosure | Experimental treatment | Weight of chemical per addition (g) per enclosure | Loading per addition (g) per enclosure | Number of additions per enclosure | Total load (g) per enclosure in 15 weeks |
| C and D | Pulse N | 910.17 | 145.63 g N | 2 | 291.26 g N |
| P | 100.72 | 20.14 g P | 2 | 40.28 g P | |
| A and E | Press N | 60.678 | 9.71 g N | 29 | 281.59 g N |
| P | 6.715 | 1.34 g P | 29 | 38.95 g P | |
| B and F | Control | unmanipulated | |||
Every Tuesday and Friday, seven 1 L water samples (~ 30 cm depth) were collected, one from each enclosure and one from the Blind Channel. These were analyzed for orthophosphate according to the methods of Stainton et al. (1977). Nitrate was measured by the ultraviolet screening method and alkalinity was measured by the titration method (APHA 1992). Seven additional samples were collected at the same time and submitted to Norwest Labs (Winnipeg) for analysis of nitrate+nitrite using the automated cadmium reduction method (APHA 1992).
Every Tuesday, three 1 L samples of water were collected from random positions in each enclosure and from the Blind Channel for analysis of phytoplankton carbon fixation rate, chlorophyll and particulate phosphorus content. The samples were collected using a water column sampler, which consisted of a transparent acrylic tube, 6.4 cm in diameter and 50 cm in length, which could be sealed at each end with a rubber stopper. The open-ended tube was lowered vertically into the water. Once submersed, first the top and then the bottom stopper was pushed into place to obtain an integrated water column sample including both phytoplankton and zooplankton. The samples were then filtered through a 100 µm pore size mesh plankton bucket to remove the zooplankton (Hann 1991). One liter of the remaining filtered water was collected from each of the three sample sites in each enclosure and transported back to the field lab for analysis. Phytoplankton productivity was measured according to the methods of Goldsborough (1993b). One mL of radiolabeled (14C) bicarbonate solution (1 µCi/mL) was added to clear glass tubes containing 25 mL of each collected water sample. The tubes were placed in a waterbath at a constant temperature of 25ºC, provided with a light source (500 µE/m2/s), and allowed to photosynthesize over a period of four hours. Blackened glass tubes containing another 25 mL of each water sample, also with 1 mL of radiolabeled bicarbonate solution added, were placed in the same water bath over the same period as controls. After the four hour incubation period, the algal cells from each sample were collected on 0.45 µm pore size glass microfiber filters (Whatman GF/C) under vacuum. The filters containing the samples were fumed over concentrated HCl to release any residual inorganic radiolabeled bicarbonate as carbon dioxide. The filters were then placed in 5 mL glass vials of liquid scintillation cocktail (Beckman ReadySafe™). Radioactivity of the samples was determined by liquid scintillation counting in a Beckman LS 3801 scintillation counter. Using these values for radioactivity, plus measurements of the pH and alkalinity of the incubation medium, the carbon fixation rate (µgC/L/h) during the incubation period was calculated for each sample.
For chlorophyll analysis, a known volume (~ 400 mL) of each remaining water sample was filtered through GF/C filters. About 1 mL of MgCO3 solution was added to the water sample during filtration to preserve the algal cells on the filters. The filters containing the phytoplankton cells were frozen for at least 24 hours to disrupt the algal membranes. The filters were then thawed, placed in 90% methanol and stored in the dark for 24 hours to allow complete extraction of chlorophyll pigments from the algal cells. Measurements of the light absorbance of the pigment extract were made at 665 nm and 750 nm (1 cm path length) for chlorophyll a and its derivatives, using a Milton-Roy Spectronic 601 spectrophotometer. One hour after acidification with 10-3 N HCl to facilitate conversion to pheophytin, the pigment extract’s absorbance was again measured at 665 nm and 750 nm. Calculation of chlorophyll concentration (µg/L) followed Marker et al. (1980).
Six colonized acrylic rods were chosen randomly from each enclosure every Thursday. The rods from each enclosure were sampled without replacement. At the time of rod removal, the water column surrounding three of the rods in each enclosure was collected for zooplankton sampling. A modified water column sampler, with notches in the center of the both the top and bottom stopper to accommodate the diameter of the rod, was carefully lowered around the rod. The stoppers were fitted into place around the rod and into each end of the sampler tube. The sampler tube and the rod were then drawn up simultaneously as one unit. The water column sample was slowly drained from the tube to minimize disturbance of the periphyton colonizing the rod and the water was collected for zooplankton sampling (Hann 1995). In the field, each rod was separated at the pre-scored notches into its predetermined subsample segments, using two pairs of needle-nosed pliers placed close to the notches to minimize hand contact with the rods. The two segments for productivity were placed in capped tubes (one clear and one blackened) in 25 mL of water from that enclosure that had previously been filtered GF/C filters. The two segments for chlorophyll analysis were placed in empty capped tubes for transport back to the field lab.
Periphyton productivity was analyzed as for phytoplankton, except that the colonized rod segment was retained on the filter and placed into scintillation cocktail with the filter. Periphyton chlorophyll analysis followed the phytoplankton procedure as above, except that the colonized rod segments were placed in the solvent.
Once the metaphyton community had developed (beginning 29 June, week 6), it was sampled every Thursday at the same time as periphyton was sampled, using the same six random locations within each enclosure as for periphyton. If metaphyton was present at a location, a fine mesh sieve (area 175 cm2) was carefully slid into place under the metaphyton and lifted straight up through the mat (Goldsborough 1993b). Any metaphyton overhanging the edges of the sieve was carefully sheared off, so that the surface area of sample obtained corresponded to the known area of the sieve. The samples were dried to constant weight at 104ºC and their dry weights (g/cm2) were recorded.
Macrophytes were sampled every second Monday using a macrophyte sampler modified from Pip and Stewart (1976). Three random locations within each enclosure were sampled. The macrophyte sampler was lowered carefully into an enclosure, enclosing the macrophytes in a 30 x 30 cm area of the bottom. The sampler consisted of a heavy-gauge aluminum frame with inside dimensions of 30 x 30 x 100 cm, which was wrapped in two layers of fine mesh screen, the outer nylon mesh screen and the inner Nitex™ screen (pore size 100 µm), around the four vertical sides to contain the zooplankton. A sharp moveable blade attached to screen was set in guide slits at the bottom of the frame with a stationary blade facing it along one side of the bottom. When a messenger released the trigger mechanism, taut heavy-duty rubber bands attached to the ends of the blades pulled the moveable blade and attached screen rapidly across the bottom to meet with the stationary blade. This effectively sheared the macrophytes just above the sediment surface, containing them and their associated epiphytes and zooplankton within the mesh-enclosed sampler. The sampler was then lifted out of the water column and placed in a rubber tray, where carbon-filtered water was used to wash the sample off the mesh screen into the tray. This water was then filtered through 100 µm pore size mesh to remove the zooplankton. The filtered water and the macrophytes were then placed in a large closed container and shaken vigorously to dislodge the epiphyton from the macrophytes.
In the field lab, macrophyte samples were placed in plastic bags and refrigerated at 4ºC prior to measurement. They were later dried to constant weight at 104ºC and weighed.
Epipelon sampling was carried out biweekly on alternate Mondays to macrophyte sampling. Samples of surface sediments (~ 1 to 2 cm deep) were obtained from three random locations in each enclosure. Lengths of open-ended PVC pipe (10.2 cm diameter) were lowered through the water column and pushed firmly into the bottom sediments to delineate the sample sites. Using a hand operated vacuum pump attached to a long piece of tubing supported by a length of acrylic rod, the surface sediments inside the diameter of the PVC pipe were aspirated into a vacuum bottle. The sediment slurry was transferred to black-walled beakers and stored in a dark drawer for 24 hours to allow the sediment to settle. The overlying water was carefully drawn off without disturbing the underlying sediment. The beakers were then transported to an outdoor site where they would receive natural irradiance for at least 18 hours. At the site, each wet sediment surface was covered by a circle of lens paper. The blackened sides of the beakers prevented light penetrance through the sides. After 18 hours, the lens paper circles were carefully removed from the beakers and placed in 100 mL of water (filtered through GF/C filters) from the same enclosure as the sediment sample. These samples were vigorously shaken for two minutes to dislodge the epipelic algae which had migrated up from the sediments into the lens paper. Subsamples of the water containing the epipelon were dispensed into clear and blackened tubes (25 mL each) for productivity analysis as described above. The remaining 50 mL subsample was filtered for chlorophyll analysis as above.
The levels of N and P in the water column of the control enclosures were low and remained relatively constant over the 15 week experimental period (Fig. 1). In the pulse enclosures, N and P levels were elevated dramatically at the time of each of the two pulse additions, although the rise was not quite so marked at the second pulse addition. Levels of N in the water column dropped off quickly and returned to pre-addition levels three weeks after the first pulse (Fig. 1A). From 10 July to 24 July, N levels in the pulse enclosures were similar to those in the control enclosures. After the second pulse addition on 25 July, N levels in the pulse enclosures dropped, within two weeks of the second pulse, to similar levels as in the press enclosures. Press N levels rose slightly at the time of the first addition on 20 June (Fig. 1B), after which they were relatively consistent throughout the remainder of the experiment. Levels of P in the water column followed a similar pattern, except that P levels did not drop back as dramatically. The second P pulse on 25 July produced a noticeably smaller peak than the first pulse. P levels in the pulse enclosures never dropped back to pre-addition conditions. Although the major peaks dropped off quickly, the P levels in the pulse enclosures showed an increasing trend through to the end of the experimental period. The press P levels increased from the first addition on 20 June through to the end of the experiment.
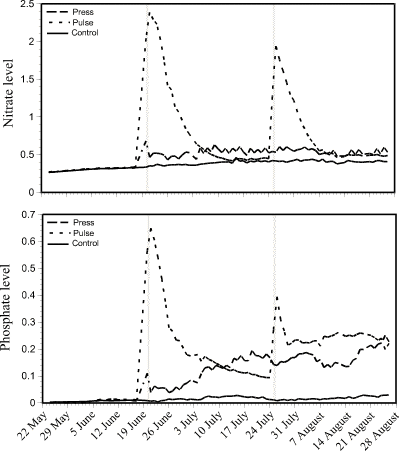
Figure 1. Nitrate (as NaNO3) levels
(A - top) and phosphoate (as NaH2PO4-2H2O)
levels (B - bottom) in the water column over a 15-week period in
press, pulse, and control treatments. Vertical bars indicate N
and P pulse additions on 20 June and 25 July 1994. Press
additions of N and P were made thrice weekly from 20 June to 24
August 1994.
Phytoplankton chlorophyll concentration was similar in all three experimental treatments (Fig. 2). Levels were highest early in the season in all enclosures, prior to nutrient addition (Fig. 2A). A peak of 20 to 30 µg/L occurred around the end of May in all enclosures. Chlorophyll concentrations then declined slowly and remained around 2 to 8 µg/L for the remainder of the summer in all enclosures. Phytoplankton productivity profiles (Fig. 2B) exhibited the same seasonal trend.
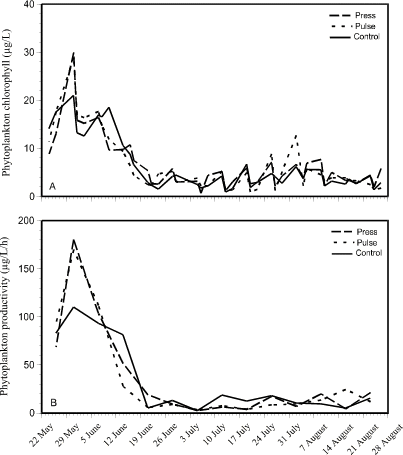
Figure 2. Changes in phytoplankton
chlorophyll concentration (A - top) and photosynthesis (B -
bottom) over a 15-week period in press, pulse, and control enclosures.
Periphyton production increased in all enclosures through the 15 week experimental period (Fig. 3). Periphyton productivity increased to 6 µgC/cm2/h in the pulse enclosures at the time of the first addition on 20 June (Fig. 3B). By 3 July, periphyton productivity in the pulse enclosures decreased to the productivity rate in the control enclosures (around 3.5 µgC/ cm2 /h), after which they both leveled off and remain nearly constant for the rest of the experimental period. There was no noticeable increase in periphyton productivity in the pulse enclosures at the time of the second pulse addition. Press treatment periphyton productivity peaked at 5.5 µgC/cm2/h somewhat later than in the pulse treatment, around 14 July. For the remainder of the season, periphyton productivity in the press enclosures remained around 4 µgC./cm2/h, slightly higher than in the pulse and control treatment enclosures. Periphyton chlorophyll concentrations (Fig. 3A) exhibited much the same trend as for productivity. Chlorophyll concentrations in pulse and control treatments were similar, while in the press treatment, chlorophyll concentrations continued increasing to the end of the experiment.
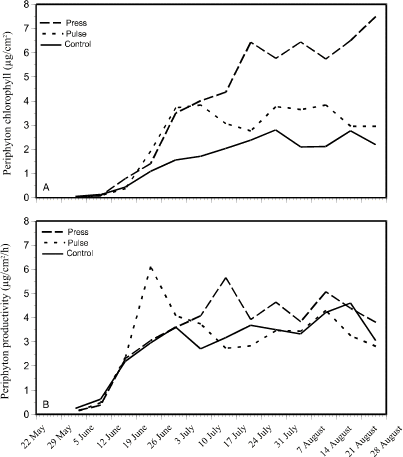
Figure 3. Changes in periphyton
chlorophyll concentration (A - top) and photosynthesis (B -
bottom) over a 15-week period in press, pulse, and control enclosures.
Epipelon chlorophyll concentrations were highest in all enclosures at the beginning of the experiment (Fig. 4). However, it should be noted that even these highest concentrations were not high as compared to periphyton concentrations (0.2 to 0.3 µg/cm2 for epipelon compared to 2 to 7.5 µg/cm2 for periphyton). By 20 June, when nutrient addition began, epipelon chlorophyll levels in all enclosures were < 0.1 µg/cm2 and remained so for the rest of the experiment (Fig. 4A). Epipelon productivity was highest (85 to 110 µgC/cm2/h) in all enclosures around 20 June (Fig. 4B) then decreased to < 20 µgC/cm2/h from 3 July to the end of the experiment.
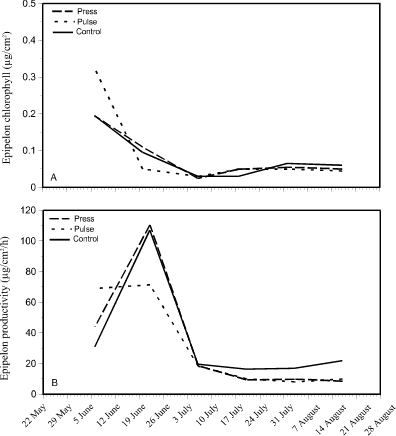
Figure 4. Changes in epipelon
chlorophyll concentration (A - top) and photosynthesis (B -
bottom) over a 15-week period in press, pulse, and control enclosures.
Metaphyton was absent from all enclosures until about a week after the first nutrient additions on 20 June (Fig. 5A). Metaphyton developed in both the press and the pulse enclosures around 3 July, after which the metaphyton biomass in both treatments increased rapidly. Both pulse and press treatments showed an increasing trend in metaphyton biomass production which continued to the end of the experimental period. Press treatment biomass was slightly higher (200 to 350 g/m2) than pulse treatment biomass (150 to 300 g/m2) from 24 July to the end of the experiment. Metaphyton was virtually absent from the control enclosures, although there is a small amount (100 g/m2) produced around mid-August.
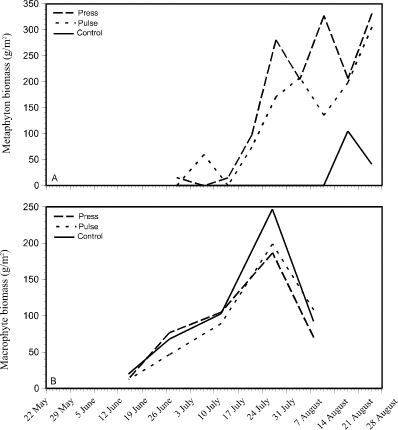
Figure 5. Changes in the dry weight of
metaphyton (A - top) and macrophytes (B - bottom) over a 15-week
period in press, pulse, and control treatments.
Macrophyte biomass showed very similar trends in all enclosures over the 15 week experimental period (Fig. 5B). Macrophyte biomass increased steadily from around 20 g/m2 in early June to 200 to 250 g/m2 at its peak around 26 July, after which biomass decreased.
The profiles of the N and P levels in the water column closely reflected the patterns of pulse and press nutrient additions. Nutrient levels in the water column of treated enclosures remained consistently higher than in the controls, except for a short period in the pulse enclosures from 10 July to 24 July, when N levels were similar to levels in the controls. This suggests that the added nutrients were remaining in the water column for a significant period of time (in terms of algal life cycles, which are measured in periods of weeks), rather than sedimenting immediately to the bottom. The peaks in N and P at the time of the second pulse additions were smaller, probably due to the presence of increased algae and macrophytes which could use nutrients from the water column quickly. The smaller peak in P at the time of the second pulse addition suggests that phosphorus was being luxury-consumed, particularly by metaphyton, periphyton and macrophytes. The increasing P levels in both the press and the pulse treatments as the season progressed suggest that more P was being added than the plants could consume. Both the N and P levels in the pulse treatments fell below the levels in the press treatments prior to the second nutrient pulse, although only for a period of about two weeks. This was reflected in both the periphyton chlorophyll and productivity profiles. Periphyton productivity in the pulse enclosures dropped as the nutrient levels dropped, while at the same time, periphyton productivity in the press enclosures was increasing steadily.
Phytoplankton appeared to have been unaffected by either pattern of nutrient addition. The early peak in phytoplankton productivity was likely the result of a normally-occurring spring phytoplankton bloom. With the onset of nutrient additions, phytoplankton appeared to have been outcompeted for the available nutrients by other algal communities. Epipelon also appeared to have been relatively unaffected by either the timing or the magnitude of nutrient addition. The increase in productivity at the time of the first nutrient addition was probably coincidental. The control showed the same increase in productivity, ruling out the nutrition addition as a causative factor. Epipelon productivity could be expected to be higher early in the season, when irradiance at the sediments was higher, before there was much macrophyte or algal biomass above to shade the sediments. The water column within all the enclosures became clear as a result of the sheltering effect of the curtain walls against strong water movements. The rise in epipelon productivity probably coincided with reduced turbidity of the water column.
It is difficult to assess whether there was a difference in response by periphyton to the timing and magnitude of nutrient additions. The trends over the season were similar in all three treatments, suggesting that the driving factor was a seasonal response to the light regime, rather than a response to manipulated nutrient levels. Periphyton productivity in the press treatment was higher, with correspondingly higher biomass, suggesting that the periphyton community gained some competitive advantage from the consistently elevated supply of N and P in the water column. While a measurement of macrophyte epiphyton is not presented here, we conjecture that its productivity profile would be similar to that of the periphyton on the acrylic rods, at least until the point when macrophytes began to decline. The high periphyton, and probably epiphyton, biomass at the time of the second nutrient pulse on 25 July would have provided ample “starter” conditions for the proliferation of metaphyton which began at that time. The timing and magnitude of the nutrient additions did not seem to have a noticeable impact on metaphyton development, as it developed equally in both press and pulse treatments. The major difference between these two treatments and the controls, which did not develop metaphyton at this time, was the higher level of phosphorus in the water column of the press and pulse enclosures. Nitrogen was also higher, but it had been at a consistently higher level in the press treatment than in the controls throughout most of the period. It was as the phosphorus levels increased in both the press and pulse treatments that metaphyton flourished. There was probably a requirement for high irradiance, as well as high nutrient availability, which would also factor into the similar timing of metaphyton development in both press and pulse treatments. There was a noticeable decline in macrophyte biomass around the time that the metaphyton started to proliferate. This may have been the result of increased shading by the metaphyton mats, or increased competition for nutrients from the metaphyton, or a combination of the two. Periphyton productivity had leveled off by the time metaphyton was developing in the treatment enclosures, but there was no evidence of a real decline in productivity.
Macrophyte biomass was a significant factor in at least two ways. Macrophytes provided abundant surface area for colonization by epiphyton, as well as initial support for large metaphyton mats. Macrophytes also competed for nutrients, both in the water column and from the sediments. Macrophytes are often thought to have a competitive advantage over algae in terms of nutrient uptake, given that they have access to nutrients in the sediments via their roots, as well as being able to absorb nutrients from the water column through their leaves. In this study, much of the added nutrient remained in the water column for extended periods, suggesting that there may not have been much competitive advantage imparted to the macrophytes in terms of increased sediment nutrients. The macrophyte biomass profiles suggest that macrophyte growth was not significantly influenced by either pattern of nutrient addition. The pulse and press profiles were similar to the control profile, suggesting that seasonal factors such as irradiance and temperature were more important in influencing their growth pattern than nutrient additions. However, the presence of high macrophyte biomass at the same time as phosphorus levels were increasing in both press and pulse enclosures (circa 25 July) was probably a factor in the development of metaphyton mats, as suggested earlier.
The control enclosures were dominated by periphyton (and probably epiphyton with high macrophyte biomass to provide abundant substrata for colonization), remaining in the open marsh state that is maintained in Delta Marsh by periodic natural disturbances. Both the press and the pulse treatment enclosures were initially dominated by periphyton, but later shifted to metaphyton dominance. This transition suggests the development of a sheltered marsh state, as predicted by the model of Goldsborough and Robinson (1996).
The transition to a sheltered marsh state in treatment enclosures supports our hypothesis that nutrient additions will alter the structuring of the algal assemblages in a wetland. However, there does not appear to be much difference in growth response to the two patterns of treatment, pulse or press. Both patterns of addition produced a similar growth response among algal communities and macrophytes. The significant difference was between the nutrient addition treatments and the control treatment. The transition to a metaphyton-dominated system under high nutrient conditions has been observed in other studies. Fong et al. (1993) found that macroalgal and cyanobacterial mats were better competitors than phytoplankton for high levels of nutrients in experimental microcosms. Murkin et al. (1994) found that metaphyton responded to periodic (press) nutrient addition later in the growing season (mid-July, early August) and suggested that it took several additions of N and P to reach levels suitable for metaphyton growth. They noted a shading effect on epiphyton and phytoplankton from the metaphyton.
Metaphyton mats, indicative of a sheltered marsh, are relatively rare in Delta Marsh, except in areas undergoing drawdown and reflooding (Goldsborough and Robinson 1996). Increased decomposition of organic matter during drawdown and the subsequent flushing of nutrients from the sediments during reflooding would provide a natural nutrient enrichment event such as we have simulated in this experiment. We conclude that the transition to a metaphyton-dominated sheltered marsh observed in this experiment is due to the nutrient enrichment of the water column.
This study was funded by an Natural Sciences and Engineering Research Council (NSERC) Summer Scholarship to RLM and an NSERC Research Grant to LGG. The Portage Country Club kindly allowed access to the study site. Assistance with sampling and analyses was provided by Mandy Lloyd, Leanne Zrum, Curt Horning and Ken Sandilands. RLM benefited from many procedural and theoretical discussions with LGG and Dr. Brenda Hann.
American Public Health Association. 1992. Standard methods for the examination of water and wastewater. 18th edition. Editors: Greenberg, A. E., Clesceri, L. S., and Eaton, A. D. Washington, D.C.
Fong, P., Donohoe, R. M. and Zedler, J. B. 1993. Competition with macroalgae and benthic cyanobacterial mats limits phytoplankton abundance in experimental microcosms. Mar. Ecol. Progr. Ser. 100: 97-102.
Goldsborough, L. G. 1991. Effects of hexazinone on water chemistry and periphyton biomass and productivity in large littoral enclosures. University Field Station (Delta Marsh) Annual Report 26: 50-62.
Goldsborough, L. G. 1993a. Responses of marsh algal communities to controlled nitrogen and phosphorus enrichment. University Field Station (Delta Marsh) Annual Report 28: 35-40.
Goldsborough, L. G. 1993b. Diatom ecology in the phyllosphere of the common duckweed (Lemna minor L.). Hydrobiologia 269/270: 463-471.
Goldsborough, L. G. and Robinson, G. G. C. 1985. Effect of an aquatic herbicide on sediment nutrient flux in a freshwater marsh. Hydrobiologia 122: 121-128.
Goldsborough, L. G. and Robinson, G. G. C. 1996. Pattern in wetlands. In: R. J. Stevenson, M. L. Bothwell and R. L. Lowe (eds.) Algal Ecology in Freshwater Benthic Ecosystems, Chapter 4. Academic Press, San Diego, California.
Goldsborough, L. G., Robinson, G. G. C. and Gurney, S. E. 1986. An enclosure/substratum system for in situ ecological studies of periphyton. Arch. Hydrobiol. Beih. 106: 373-393.
Hann, B. J. 1991. Invertebrate grazer-periphyton interactions in a eutrophic marsh pond. Freshwater Biology 26: 87-96.
Hann, B. J. 1995. Response of aquatic invertebrates to experimental nutrient enrichment of a wetland. University Field Station (Delta Marsh) Annual Report, this volume.
Kusler, J. A., Mitsch, W. J. and Larson, J. S. 1994. Wetlands. Sci. Am. Jan. 1994. pp. 64-70.
Lane, R. K. 1978. Plate 18. Large Lakes in: Hydrological Atlas of Canada. Surveys and Mapping Branch, Department of Energy, Mines and Resources, Ottawa, Ontario.
Manny, B. A., Johnson, W. C. and Wetzel, R. G. 1994. Nutrient additions by waterfowl to lakes and reservoirs - predicting their effects on productivity and water quality. Hydrobiologia 280: 121-132.
Marker, A. F. H., Crowther, C. A. and Gunn, R. J. M. 1980. Methanol and acetone solvents for estimating chlorophyll a and phaeopigments by spectrophotometry. Arch. Hydrobiol. Beih. 14: 52-69.
Millar, J. B. 1989. Perspectives on the status of Canadian prairie wetlands. In: R. R. Sharitz and J. W. Gibbons (eds.) Freshwater Wetlands and Wildlife. DOE Symposium Series No. 61. pp. 829-852.
Mitsch, D. F. and Gosselink, J. G. 1993. Wetlands, Second Edition. Van Nostrand Reinhold, New York.
Murkin, H. R., Pollard, J. B., Stainton, M. P., Boughen, J. A. and Titman, R. D. 1994. Nutrient additions to wetlands in the Interlake region of Manitoba, Canada: effects of periodic additions throughout the growing season. Hydrobiologia 279/280: 483-495.
Murkin, H. R., Stainton, M. P., Boughen, J. A., Pollard, J. B. and Titman, R. D. 1991. Nutrient status of wetlands in the Interlake Region of Manitoba, Canada. Wetlands 11: 105-122.
Pip, E. and Stewart, J. M. 1976. The dynamics of two aquatic plant-snail associations. C. J. Zool. 54: 1192-1205.
Rubec, C. D. A. and Rump, P. C. 1985. Prime wetland land use monitoring activities and needs in southern Canada. Agricultural Land Drainage, Proceedings of the Third Annual Western Provincial Conference - Rationalization of Water and Soil Research and Management, Winnipeg, Manitoba, November 20-22, 1984. Manitoba Water Resources Branch, Winnipeg, Manitoba, Canada, pp. 405-429.
Schindler, D. W. 1974. Eutrophication and recovery in experimental lakes: Implications for lake management. Science 184: 897-899.
Schindler, D. W., Armstrong, F. A. J., Holmgren, S. K. and Brunskill, G. J. 1971. Eutrophication of Lake 227, Experimental Lakes Area, Northwestern Ontario, by addition of phosphate and nitrate. J. Fish. Res. Bd Can. 28: 1763-1782.
Stainton, M. P., Capel, M. J. and Armstrong, F. A. J. 1977. The chemical analysis of fresh water. Second Edition. Fisheries and Environment Canada. Miscellaneous Special Publication No. 25.
Tilman, D. 1977. Resource competition between planktonic algae: an experimental and theoretical approach. Ecology 58: 338-348.
Wetzel, R. G. 1964. A comparative study of the primary productivity of higher aquatic plants, periphyton, and phytoplankton in a large, shallow lake. Int. Rev. ges. Hydrobiol. 49: 1-61.
Wetzel, R. G. 1983. Limnology, Second Edition. Saunders College Publishing, New York.
Zoltai, S. C. 1988. Wetland environments and classification. In: Wetlands of Canada, Ecological Land Classification Series No. 24. pp. 1-26. Environment Canada, Ottawa.