D. Glen McMaster
Department of Zoology, University of Manitoba
Winnipeg, Manitoba, Canada R3T 2N2
E-mail:
|
Does host-egg removal increase the hatching success of parasitic cowbird eggs?
D. Glen McMaster Department of Zoology, University of Manitoba Winnipeg, Manitoba, Canada R3T 2N2 E-mail: |
|
Avian brood parasites lay their eggs in the nests of other birds, the hosts, which then incubate and raise the young (Payne 1977). Brood parasitism often reduces the number of host fledglings produced through damage or removal of host eggs, eviction of host eggs and/or young, and nestling competition (Payne 1977). The removal of host eggs from nests by female brood parasites has been documented in many parasitic species world-wide. Parasitic cuckoos (Cuculidae), honey guides (Indicatoridae), two finch genera (Vidua and Anomalospiza), and cowbirds (Icterinae) are known to remove or damage host eggs (Friedmann 1960, 1963; Payne 1977; Sealy 1992). In cavity-nesting species where nest sites may be limiting, egg removal may be employed as an usurpation tactic designed to force resident females to desert their nests (Lombardo et al. 1989). Female parasites usually remove host eggs in association with the laying of their own egg in the host nest (Sealy 1992), although one instance of egg removal by a male has been observed (Lombardo et al. 1989). While host-egg removal decreases host reproductive success by reducing the number of host offspring that can be raised, this behaviour may benefit the parasite and ultimately increase its reproductive success (Rothstein 1990).
Female Brown-headed Cowbirds (Molothrus ater) remove eggs from nests of many of the host species parasitized (Friedmann 1963), but removal varies in terms of number and timing. In fact host eggs are not removed from all parasitized nests but, on the other hand, one or more eggs are often removed (Mayfield 1961; Zimmerman 1963; Sealy 1992). Female cowbirds have been observed removing eggs on the day before, the same day, and the day after laying their own egg (Hann 1937; Norris 1944; Nolan 1978). When removing a host egg, cowbirds usually puncture it with their bill, carry it away, and sometimes eat it (Hann 1937; Scott et al. 1992; Sealy 1992).
Several hypotheses have been formulated to explain the function of host-egg removal behaviour (review in Sealy 1992). Enhancing the incubation efficiency of the host is one explanation given for the function of this behaviour. The incubation capacity of the host is limited by both the amount of heat that can be produced physiologically, and by the number of eggs that can physically contact the brood patch. The addition of a parasitic egg to a host clutch increases both the number and volume of eggs a host must incubate. This increase in clutch size could result in inefficient host incubation that may jeopardize the parasitic egg's chances of hatch successfully (Davies and Brooke 1988). Therefore, the incubation efficiency hypothesis states that host-egg removal by female brood parasites reduces host clutch volume to within the limits of the host's incubation capacity, optimizing the hatching success of the cowbird egg.
In 1992 and 1993, I located Yellow Warbler (Dendroica petechia) nests in the forested dune-ridge at Delta Marsh (50°11'N, 98°19'W) on the properties of the University of Manitoba Field Station and Portage Country Club. I monitored nests found before clutch initiation until the first egg was laid (Laying Day 1 = LD1). I assigned at random one of three treatment groups to each nest on LD1 to examine possible manipulation of host incubation behaviour by cowbirds. Treatment 1 consisted of switching a single, viable cowbird egg for a single host egg. In Treatment 2, I also added a single cowbird egg, but in this experiment, I did not remove a host egg. Treatment 3 was a control where cowbird eggs were neither added nor host eggs removed. Most cowbird eggs were added on the morning of LD1 (0700-1100h C.S.T.), although several were added later on the afternoon of LD1 (1200-1300h C.S.T.).
Viable cowbird eggs were obtained from naturally parasitized nests of 6 different species. An effort was made to use only cowbird eggs of known age, that had been exposed to minimal incubation (i.e. had been found during early egg-laying, or preferably even before the host had initiated its clutch).
An effort was made to minimize the amount of time a cowbird egg spent out of a natural nest. Any cowbird eggs found on a given day were added to a Yellow Warbler clutch that same day if possible. If no warbler clutches were available for parasitism eggs were kept overnight in a cardboard egg crate in a refrigerator at 10°C. Cool temperatures arrest embryonic development and allow the embryo to remain viable for much longer periods than would normally be possible (Drent 1975).
Five eggs is the modal Yellow Warbler clutch size at Delta Marsh (Sealy 1992). However, 4-egg clutches are also common, especially later in the breeding season. At the moment of the experimental parasitism on LD1 there is no way to predict whether the female will eventually lay 4 or 5 eggs. Therefore, in order to keep the number of host eggs constant between nests, once clutches were complete host eggs were transferred to 4-egg clutches to ensure all nests had 5 eggs. As 5-egg females may be able to incubate larger clutches more efficiently than 4-egg females, the influence of natural clutch size was investigated using the General Linear Models procedure.
In 1992 once the clutch was complete, I measured all eggs, and calculated egg volume using the formula V=kLB2 (k is a constant, k=0.515 for cowbird eggs and k=0.497 for Yellow Warbler eggs; L=egg length; B=egg breadth) (Hoyt 1979; Mills 1987).
Nests were revisited 1-2 days before they were predicted to begin hatching. When possible, the identity (cowbird or warbler) of the first egg to hatch was noted. In nests where cowbird eggs did not hatch, hatching order was not recorded as only Yellow Warbler eggs would be available to hatch first. The incubation period was defined as the day after the last host egg was laid (the first day of full incubation) up until the first egg in the clutch hatched. Therefore a single incubation period was recorded for the clutch as a whole. Hatching spread was defined as the date from the first egg hatching to the day the last egg hatched.
In 1992 a total of 185 Yellow Warbler nests was found, of these 103 were experimentally parasitized with live cowbird eggs. Most clutches were initiated in early June, while the latest clutch initiated was on July 4. Another 37 nests were left unparasitized and monitored as control nests. In 1993 a total of 255 Yellow Warbler nests was found, of these nests 86 were experimentally parasitized with live cowbird eggs. Another 28 nests were left unparasitized and monitored as Control nests. The majority of clutches were initiated in late May or early June, while the latest clutch initiated was on June 29.
Mean ambient temperatures at Delta Marsh for the months of June and July 1992 were cooler than the average temperature for 1951-1980. The mean temperature for June 1992 was 14.88°C compared to 16.5°C ± 1.6 (SD) for the 1951-1980 average (Environment Canada data). The mean temperature for July 1992 was 15.13°C compared to 19.1°C ± 1.2 (SD) for the 1951-1980 average (Environment Canada data). Mean daily temperature at Delta Marsh for the month of June 1993 was slightly cooler (15.7°C) than the average mean daily temperature for the period of 1951-1980 (16.5°C ± 1.6 SD) (Environment Canada data). The mean daily maximum (21.4°C) and minimum (10.03°C) for June 1993 were also only slightly cooler than those from 1967-1990 (mean daily maximum = 22.3°C, mean daily minimum = 10.9°2C).
Eggs were not measured in 1993, however 1992 data showed that clutch volume differed significantly between treatments (GLM, ANOVA, F = 60.58, p < 0.0001, N = 69). Treatment 2 clutches (mean = 9.91 mL, N = 18) were larger than Treatment 1 clutches (mean = 8.57 mL, N = 29), which in turn were larger than Control clutches (mean = 7.17 mL, N = 22).
The majority of cowbird eggs that hatched in both 1992/1993 combined did so before (45%, N = 23) or the same day as Yellow Warbler eggs hatched (41%, N = 21). However treatment had no significant effect on the hatching order of parasitized clutches in 1992 (c2 = 2.87, p = 0.238), 1993 (c2 = 0.77, p > 0.6), or 1992/1993 combined (c2 = 1.42, p > 0.4). Therefore, the likelihood of a cowbird egg hatching before warbler eggs did not differ between Treatment 1 and Treatment 2 clutches.
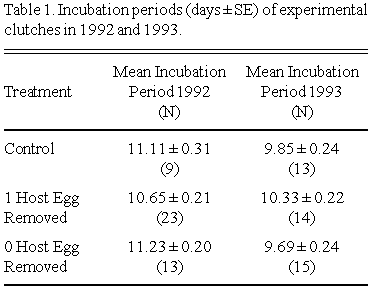
In 1992 the incubation period of a clutch was not significantly affected by either treatment (GLM, ANOVA, F = 2.16, p = 0.13, N = 39) or natural clutch size (GLM, ANOVA, F = 0.34, p = 0.56, N = 39) (see Table 1). In 1993, however, although treatment did not affect incubation period (GLM, ANOVA, F = 2.19, p = 0.31, N = 42) (Table 1), the effect of natural clutch size approached significance (GLM, ANOVA, F = 3.86, p = 0.057, N = 42) due to a shorter incubation period for 5-egg clutch females (mean = 9.69 days) than 4-egg clutch females (mean = 10.22 days). However, for 1992/1993 combined, neither natural clutch size (GLM, ANOVA, F = 0.82, p = 0.45, N = 85) nor treatment (GLM, ANOVA, F = 0.04, p = 0.96, N = 85) had a significant effect on the length of the incubation period.
Mean hatching spreads of experimental clutches in 1992 and 1993 are presented in Table 2. Neither treatment nor natural clutch size significantly influenced hatching spread in 1992 (GLM, ANOVA, F = 1.18, p = 0.344, N = 34), in 1993 (GLM, ANOVA, F = 0.22, p = 0.95, N = 39) or in 1992/1993 combined (GLM, ANOVA, F = 0.36, p = 0.94, N = 76).
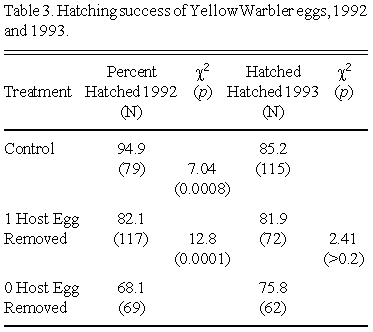
In 1992 Yellow Warbler hatching success differed significantly between Control, Treatment 1 and Treatment 2 clutches (c2 = 18.17, p = 0.0001). Host-egg hatching success was significantly higher in Control than Treatment 1 clutches, which in turn experienced significantly higher hatching success than Treatment 2 clutches (see Table 3). However, Yellow Warbler hatching success in 1993 did not differ significantly between treatments (c2 = 2.41, p > 0.20)(Table 3).
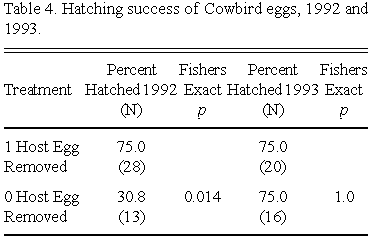
Cowbird egg hatching success in 1992 differed significantly between Treatment 1 and 2 clutches (Fishers Exact Test, p = 0.017, two-tailed). Cowbird eggs in Treatment 1 clutches experienced significantly greater hatching success than did eggs in Treatment 2 clutches (Table 4). However in 1993 treatment had no significant effect on cowbird egg hatching success (Fishers Exact Test, p = 1.0, two-tailed) (Table 4).
In 1992 the removal of a single host egg on the day of parasitism had a significant impact on the hatching success of both cowbird and host eggs. Cowbird and Yellow Warbler eggs experienced significantly higher hatching success when a host egg was removed from the clutch than when no host egg was removed; findings consistent with Davies and Brooke's (1988) hypothesis. However, host egg removal in 1993 did not have a significant effect on the hatching success of either cowbird or host eggs. The discrepancy between the hatching success results of 1992 and 1993 could be explained by differences in ambient temperature during incubation between years. 1992 was a cooler summer than 1993 and was also cooler than the 30 year average at Delta Marsh. Perhaps cooler temperatures in 1992 made it more difficult for female Yellow Warblers to incubate large clutches, resulting in lower hatching success in large clutches.
Female cowbirds may derive a number of benefits from host-egg removal behaviour. Female cowbirds have been observed eating host eggs they have removed from the nest (Hann 1937; Norris 1944; Mayfield 1960; Nolan 1978; Scott et al. 1992). An egg could represent a valuable source of nutrition for a cowbird, as well as providing a source of calcium if the egg-shell is eaten (see Sealy 1992). Female cowbirds may require considerable calcium for egg-shell formation as they lay such large clutches (Ankney and Scott 1980). However, the frequency with which eggs are removed and not eaten by female cowbirds suggests the primary purpose of egg removal is not simply for food alone (Scott et al. 1992).
By removing a host egg, the female cowbird may reduce crowding and competition in the nest once the cowbird chick hatches (Mayfield 1960; Blankespoor et al. 1982). Parasitic nestlings hatching in the nests of other bird species must compete with host nestlings for food brought to the nest by the parents. Newly hatched nestlings of some parasitic species kill their host nestmates (honeyguides and Tapera naevia cuckoo) or eject them from the nest (cuckoos), thereby reducing the competition they must face (Livesey 1939; Hamilton and Orians 1965; Morton and Farabaugh 1979; Wyllie 1975; Payne 1977). Cowbird nestlings do not attack host young, instead they often crowd and outcompete host young for food (Friedmann 1963; Rothstein 1990). As cowbird nestling weight has been shown to be inversely related to the number of competitors in the nest (Hatch 1983), removal of a host egg could improve the cowbird chick's chances of survival. By removing a host egg early in the egg-laying period the female cowbird could also increase the chances the cowbird chick will hatch first, by eliminating a host embryo that may have already undergone some development (Berger 1951).
Due to conflicting results from the two summers, no firm conclusions may be made about the function of host-egg removal with regard to the incubation efficiency hypothesis. Hatching success experiments will be repeated again in 1994, in an attempt to provide clear evidence either for or against the incubation efficiency hypothesis.
I thank D. Beattie, K. Caldwell, D. Froese, S. Gill, G. Goulet, P. Grieef, P. Miller, D. Neudorf, and L. Zdrill for assistance in the field. I wish to thank Dr. Spencer G. Sealy who has guided me throughout this study. Thanks to the staff at Delta Marsh for their hospitality, and to the officers of the Portage Country Club for allowing me to conduct research on their property. Statistical advice was provided by the University of Manitoba Statistical Advisory Service. This project was funded by an NSERC Postgraduate Scholarship, a University of Manitoba Fellowship, and a Manitoba Naturalists Society Scholarship to myself, and an NSERC Research Grant to Dr. S. G. Sealy.
Ankney, C. D., and Scott, D. M. 1980. Changes in nutrient reserves and diet of breeding Brown-headed Cowbirds. Auk 97: 684-696.
Berger, A. J. 1951. The cowbird and certain host species in Michigan. Wilson Bull. 63: 26-34.
Blankespoor, G. W., Oolman, J., and Uthe, C. 1982. Eggshell strength and cowbird parasitism of Red-winged Blackbirds. Auk 99:363-365.
Davies, N. B., and Brooke, M. de L. 1988. Cuckoos versus reed warblers: adaptations and counteradaptations. Anim. Behav. 36: 262-284.
Drent, R. 1975. Incubation. In Farner, D. S., King, J. R., and Parkes, K. C. (eds.), Avian Biology, Vol. 5: 333-420. New York: Academic Press.
Environment Canada, Atmospheric Environment Service. Canadian Climate Normals, Temperature and Precipitation 1951-1980. Prairie Provinces.
Friedmann, H. 1960. The parasitic weaverbirds. U.S. Natl. Mus. Bull. 223: 1-196.
Friedmann, H. 1963. Host relations of the parasitic cowbirds. U.S. Natl. Mus. Bull. 233: 1-276.
Hamilton, W. J., and Orians, G. H. 1965. Evolution of brood parasitism in altricial birds. Condor 67: 361-382.
Hann, H. W. 1937. Life history of the oven-bird in southern Michigan. Wilson Bull. 49: 145-237.
Hatch, S. A. 1983. Nestling growth and relationships of Brown-headed Cowbirds and Dickcissels. Wilson Bull. 95: 669-671.
Hoyt, D. F. 1979. Practical methods of estimating volume and fresh weight of bird eggs. Auk 96: 73-77.
Livesey, T. R. 1939. Egg stealing by Khasia Hills Cuckoo (Cuculus canorus bakeri Hartert). J. Bombay Nat. Hist. Soc. 40: 561-564.
Lombardo, M. P., Power, H. W., Stouffer, P. C., Romagnamo, L. C., and Hoffenberg, A. S. 1989. Egg removal and intraspecific brood parasitism in the European Starling (Sturnus vulgaris). Behav. Ecol. Sociobiol. 24: 217-223.
Mayfield, H. F. 1960. The Kirtland's Warbler. Cranbrook Institute of Science. Bull. 40: 1-242.
Mayfield, H. F. 1961. Vestiges of proprietary interest in nests by the Brown-headed Cowbird parasitizing the Kirtland's Warbler. Auk 78: 162-166.
Mills, A. M. 1987. Size of host egg and egg size in Brown-headed Cowbirds. Wilson Bull. 99: 490-491.
Morton, E. S., and Farabaugh, S. M. 1979. Infanticide and other adaptations of the nestling Striped Cuckoo Tapera naevia. Ibis 121: 212-213.
Nolan, V. Jr. 1978. The ecology and behavior of the Prairie Warbler Dendroica discolor. Ornithol. Monogr. 26: 1-595.
Norris, R. T. 1944. Notes on a cowbird parasitizing a Song Sparrow. Wilson Bull. 56: 129-132.
Payne, R. B. 1977. The ecology of brood parasitism in birds. Ann. Rev. Ecol. Syst. 8: 1-28.
Rothstein, S. I. 1990. A model system for coevolution: avian brood parasitism. Annu. Rev. Ecol. Syst. 21: 481-505.
Scott, D. M., Weatherhead, P. J., and Ankney, C. D. 1992. Egg-eating by female Brown-headed Cowbirds. Condor 94: 579-584.
Sealy, S. G. 1992. Removal of Yellow Warbler eggs in association with cowbird parasitism. Condor 94: 40-54.
Wyllie, I. 1975. Study of cuckoos and Reed Warblers. Br. Birds 68: 369-378.
Zimmerman, J. L. 1963. A nesting study of the catbird in southern Michigan. Jack-Pine Warbler 41: 142-160.
{kind=link}
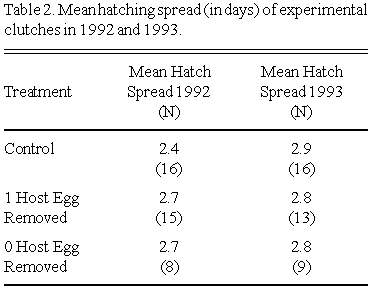{kind=link}
{kind=link}
{kind=link}