Norm C. Kenkel
Department of Botany, University of Manitoba
Winnipeg, Manitoba, Canada R3T 2N2
E-mail: kenkel@ccu.umanitoba.ca
|
Response of whitetop meadow vegetation to disturbance: a five-year experiment
Norm C. Kenkel Department of Botany, University of Manitoba Winnipeg, Manitoba, Canada R3T 2N2 E-mail: kenkel@ccu.umanitoba.ca |
|
In prairie marsh ecosystems, disturbance plays an important role in the maintenance of floristic and landscape diversity. At the scale of the whole marsh, water level fluctuations are of critical importance (van der Valk and Davis 1978; van der Valk 1981). The prairie regions of North America are characterized by alternating cycles of high and low annual precipitation, which in turn results in a cycle of alternating flooding and drawdown periods in prairie wetlands. High water levels in the marsh kills emergent macrophyte vegetation, but vegetation regeneration from the seed bank and surviving underground parts occurs when water levels fall again. Van der Valk hypothesized that such periodic disturbance is essential to the regeneration of prairie wetlands. In recent years, concerns have been raised that anthropogenic control of water levels in wetlands may permit highly competitive species to competitively exclude less competitive ones, resulting in lowered species and habitat diversity (Kenkel 1993).
More localized disturbances may also be important in maintaining species and habitat diversity in prairie wetlands. Examples included muskrat 'eat outs' of macrophyte vegetation, and the 'trampling' effects on vegetation by deer and humans. While the effect of such localized disturbances on prairie marsh vegetation has not been extensively studied, they may be very important. Prairie marshes are characterized by a few dominant emergent macrophytes that, through clonal growth, form extensive monodominant stands. Associated 'understory' species in such stands are generally opportunistic, non-clonal annuals and short-lived perennials. These 'understory' species usually occur at low frequency and abundance, but may be locally abundant in small disturbed patches within an extensive monodominant stand.
An example of the possible importance of small-scale disturbance in whitetop meadows is given by Neill (1990a,b). While his study was designed to examine the effect of nutrient addition (N and P) on emergent macrophyte productivity, an important secondary effect related to local disturbance was observed. The author found that nitrogen limited growth of both Scolochloa festucacea and Typha x glauca at the Delta Marsh, Manitoba. In whitetop meadows, nitrogen significantly increased productivity in the year following application. However, a quite different effect was observed in the second year, as whitetop biomass decreased dramatically while biomass of other species (particularly the annual Atriplex patula) showed a dramatic increase. Neill hypothesized that whitetop growth in the second year was inhibited by a mat of fallen litter from the previous year. Thus a disturbance (in this case, nutrient addition) caused a 'mulching' effect of litter that in turn resulted in a localized increase in species diversity. Neill also hypothesized that this effect would be short-lived, as whitetop was expected to quickly spread back into these 'mulched' patches through vegetative propagation. This was not verified, however, as his experiment was terminated after two years.
In this study, I investigate the long-term effects of small-scale patch disturbance on productivity and species diversity in a whitetop meadow at Delta, Manitoba. I used different levels of salt (sodium chloride, NaCl) addition to 'simulate' a gradient of increasing disturbance in a controlled, replicated experiment.
The Delta Marsh, a large (21,870 ha) lacustrine prairie wetland at the south end of Lake Manitoba, is one of the largest remaining natural marshes on the North American prairie. The marsh is connected to Lake Manitoba by a series of channels crossing a sandy dune ridge. The dominant macrophyte species in the marsh include Scirpus sp., Typha sp. (mainly the hybrid cattail T. glauca, T. angustifolia x T. latifolia), Carex atherodes, Scolochloa festucacea, and Phragmites australis. A complete description of the vegetation is given by Löve and Löve (1954) and Walker (1965). Soils are variable, but are predominantly silt loams, sand loams, and silty clay loams overlain by a poorly to moderately decomposed organic layer. The substrate is somewhat basic, and generally slightly to moderately saline. Water levels in the marsh typically drop over the growing season, typically between 20 and 50 cm.
The study site used in the experiment is located on the property of the University of Manitoba Field Station (50°11'N, 98°19'W, elevation » 250 m), just south of the main lodge. The site is a vegetatively homogeneous wet meadow dominated by whitetop (Scolochloa festucacaea). Associated 'understory' species, which occur at much lower abundance, include sow thistle (Sonchus arvensis), Canada thistle (Cirsium arvense), smartweeds (Polygonum sp.), and various mints (mainly Teucrium occidentale, but also Mentha arvensis, Lycopus asper and Stachys palustris). Opportunistic annual species (mainly Chenopodium rubrum, Atriplex patula and Aster simplex) occur on exposed substrates.
Whitetop (Scolochloa festucacea) is a rhizomatous, emergent hydrophytic grass. It is especially common in Manitoba, Saskatchewan and North Dakota, where it occurs in seasonally flooded meadows. It may be found at the centers of small basins (prairie 'potholes'), and along the shallow edges of larger marshes in areas inundated by snowmelt water. The species tolerates a wide range of conductivities, but does best in moderately saline habitats (Neill 1990a,b). On the prairies, vernal growth is initiated between mid-April and mid-May. Smith (1973) suggests that litter accumulation is detrimental to whitetop productivity, since burning or mowing after a season without disturbance results in an increase in productivity. He suggests that Carex atherodes will eventually outcompete whitetop in areas where significant amounts of litter accumulate.
In the fall of 1988, a 22 x 22 m study area was delineated within a homogenous whitetop meadow. Elevation varied slightly within the study area, with the north side of the region being the least flooded. Elevation dropped by 7 cm between the NE and NW corners, by 34 cm between the NE and SE corners, and by 27 cm between the NE and SW corners. To effectively block for this variation in elevation (which affects duration of flooding), a 6 x 6 latin square experimental design was established within the study area. The design consisted of 2 x 2 m plots spaced 2 m apart (Fig. 1). Between June 14 - 16, 1989 each plot was lined with 135 µm polyethylene film to a depth of 20 cm.
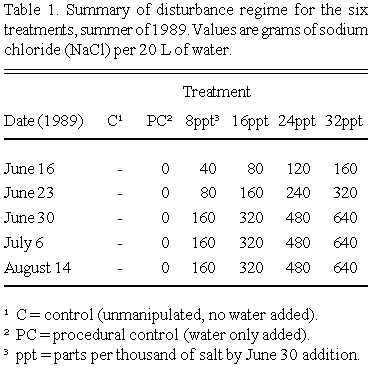
Different levels of disturbance were simulated by adding saline (NaCl) water, of varying concentrations, to the plots. There were six treatment levels: (1) control, unmanipulated; (2) procedural control, water only added; (3) 8 ppt saline water; (4) 16 ppt saline water; (5) 24 ppt saline water; (6) 32 ppt saline water. Salt (commercially available sodium chloride, Sifto® natural/no additive 'pickling salt') was dissolved in 20 L of wellwater and carefully added to each treatment plot using a siphon hose. An attempt was made to add the solution directly to the soil. Following salt addition, a few liters of water was used to carefully 'wash' any salt solution from plant leaves.
Salt solutions were added during the mid-summer of 1989, after water levels had dropped to the point that plots were no longer flooded. By this time, aboveground vegetative growth of whitetop was largely completed (Smith 1973). The salinity levels, while not high enough to kill the whitetop, were chosen so as to have a demonstrable negative effect on whitetop productivity. The highest salt level was well below that of halophyte-dominated salt pans in the vicinity. Salts were incrementally added over a two month period (June 14 - August 14, 1989), as summarized in Table 1.
A soil core (4 cm diameter, 15 cm deep) was taken from the center of each plot on June 7, 1989 (prior to the treatment applications) to measure soil pH, conductivity and percent water content. Additional soil cores were taken on July 5, 21 and 26 and August 16, 1989 to monitor changes in conductivity during 1989 (when salts were added), and on July 5, 1990 and August 12, 1993 to monitor conductivity and percent water content.
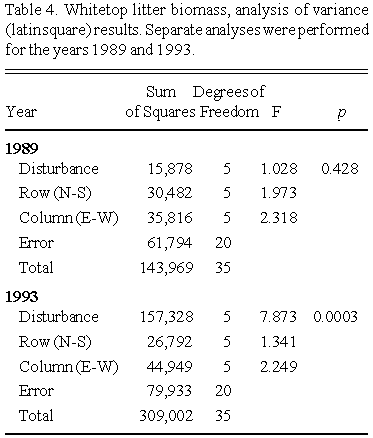
Aboveground biomass was harvested in 1989 (August 14 - 18), 1990 (August 21 - 23), and 1993 (August 3 - 5). In 1989 and 1990, harvesting was done in four 35 x 35 cm subplots within the central portion of each 2 x 2 treatment plot. All plants rooted within each subplot were harvested. Different subplots were used in the two years. In 1993, all plant material rooted within the central 1 x 1 m portion of each plot was harvested. Aboveground biomass was expressed on a per meter square basis. Harvested plant material was sorted into species, dried at 80°C, and massed to the nearest gram. Whitetop litter was harvested in 1989 and 1993.
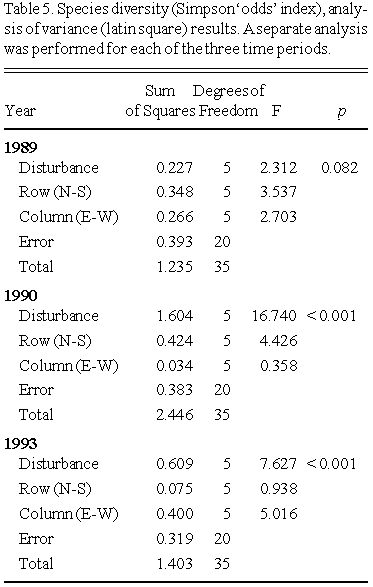
Community species diversity for each plot was measured based on the familiar Simpson's index (1- S), where:
S = S pi²
and pi is the proportion of total biomass accounted for by species i in a given plot. In this study, a variant of S known as the 'odds' index (Kvalseth 1991) was used:
K = (1/S) - 1
Analysis of variance (ANOVA, latin square model) was used to compare whitetop biomass, and community diversity, between treatments. Separate analyses were conducted for each of the three harvest years. Graphical presentation of the results used boxplots, which summarize the data in terms of the median and ranges.
Soil Factors
Soils were slightly basic in all plots. For the July 21, 1989 sample, the mean pH = 8.121 (range 7.66 - 8.36). Values for the August 6, 1993 sample were similar (mean = 8.07, range 7.72 - 8.39). No significant differences in soil pH were found between the experimental treatment levels.
Percent water content of the soil was relatively high, reflecting the relatively high organic content of the soils and the proximity of the water table in these wet meadow sites. Values in the summer of 1989 were: July 5, 1989: mean = 58% (range 38 - 65%), and August 16, 1989: mean = 55% (range 37 - 60%). Values in 1990 were somewhat higher (July 5, 1990: mean = 64%, range 58 - 72%), reflecting the wetter spring and summer weather in 1990. In 1993, values were similar to those of 1989 (August 6, 1993: mean = 56%, range 43 - 62%). No significant differences in soil water content were found between the experimental treatment levels for any of these dates.
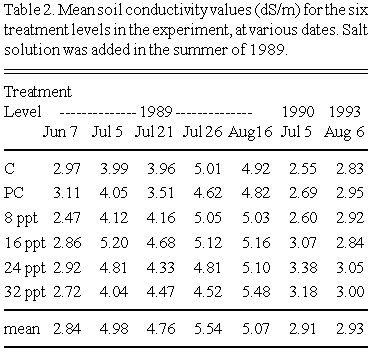
Soil conductivity trends over time are summarized in Table 2. During the summer of 1989, when salt solution was added to the plots, soil conductivity increased over the course of the summer. However, this increase occurred over all plots regardless of whether they received salt solution. The summer of 1989 was unusually hot and dry in southern Manitoba. In prairie marshes, salts in the soil may move to the substrate surface during drought as water is drawn to the surface through evapotranspirative processes. There were no significant differences in soil conductivity between treatments at any time in 1989, though values tended to be slightly higher in the high salt treatments. I hypothesize that the salt solution percolated to soil depths greater than 10 cm soon after application, and were therefore not detected by the soil cores. That the salt solution was effective in 'disturbing' the environment is evidenced by the productivity result (see below).
Conductivity values on July 5, 1990 were similar to those of June 7, 1989. This was a more typical cooler and somewhat wetter year. In 1990 soil conductivity values were highest in the most 'disturbed' plots, though this difference was not statistically significant. Values on August 6, 1993 were similar to those of 1990 and June of 1989. Note that 1993 was an unusually cool and wet summer. Even in August, the soil was saturated. As in the other time periods, there were no significant differences between treatment levels.
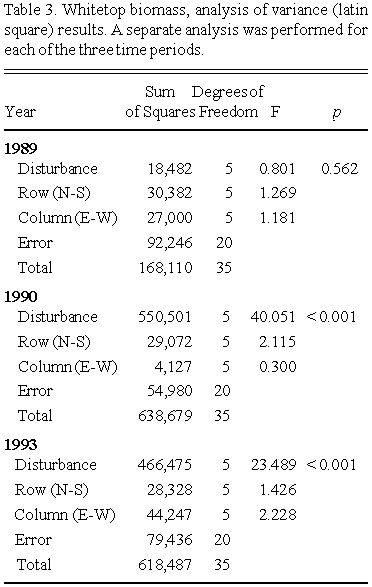
Whitetop aboveground biomass values (years 1989, 1990 and 1993) across treatments are summarized as boxplots in Fig. 2, and the corresponding ANOVA (latin square design) results are presented in Table 3.
For the 1989 harvest, no significant differences in whitetop aboveground biomass were observed between treatments. Salt solution was added in the summer of 1989, but not until the majority of the aboveground biomass had already been produced (in whitetop, shoot emergence occurs in late April, and a full developed shoot system is established by mid-June; Smith 1973). There was no significant difference in whitetop litter biomass between treatments (Table 4), but there was a significant inverse relationship between whitetop litter and aboveground biomass in the 36 plots (Fig. 3).
A statistically significant decrease in aboveground biomass with increasing salinity was seen in 1990. This probably reflects the effect of salinity on disruption of physiological processes related to rhizome production. As in many temperate species, whitetop stores photosynthates in the rhizome/root system in the fall for use the following spring. Rhizome production in whitetop occurs in August and September (Smith 1973), which is after the addition of salt solution in 1989. Salinity 'stress' likely limited rhizome production in 1989, which had a carry-over effect in the 1990 growing season. However, it should be noted that the salinity levels used in this experiment, while high enough to reduce whitetop biomass, did not lead to local (plot) extinction of the species - it was present in all plots. Also, the experimental results demonstrate that there is no difference between the two control treatments. This shows that the addition of water alone (a procedural effect) had no influence on biomass accumulation in whitetop.
By 1993, the plots receiving the highest salinities had significantly greater aboveground biomass compared to the control plots. Litter biomass was significantly less in the disturbed (higher salinity) plots (Fig. 4). As in 1989, there was a significant inverse relationship between whitetop litter and aboveground biomass (Table 4, Fig. 5).
Species diversity values (years 1989, 1990 and 1993) across treatments are summarized as boxplots in Fig. 6, and the corresponding ANOVA (latin square design) results are presented in Table 5.
No significant differences between treatment levels were seen, but the plots were quite variable. Species composition of plots also varied.
A decrease in whitetop biomass at higher salinities, combined with an increase in the abundance of 'understory' species (particularly the opportunistic annual Atriplex patula), resulted in a statistically significant increase in species diversity with increasing salinity by 1990.
In the control plots, species diversity in 1990 was lower than in 1989. This was attributable both to a decrease in 'understory' species biomass, and an increase in whitetop biomass, in 1990. The study area was flooded for a considerable period in the spring and early summer 1990, which may have inhibited germination and establishment of 'understory' species while favouring whitetop growth (Neill 1990 a,b).
In 1993, species diversity was significantly lower in the higher salinity (more highly disturbed) treatments. This was the result of both higher biomass of whitetop, and lower biomass of 'understory' species, in the high salinity plots. Higher whitetop biomass apparently inhibits the establishment and growth of 'understory' species.
This study offers strong empirical support for the notion that long-term environmental studies are necessary to obtain a complete understanding of ecological patterns and processes occurring in nature. Long-term experiments are particularly important in perturbation studies, since the short-term effects of ecosystem disturbance may differ fundamentally from effects in the longer term. In this study, the short-term effect of disturbance (1990 results) was to reduce aboveground biomass of whitetop, while increasing biomass of ruderal species. This is the 'classical' disturbance-response effect seen in many communities, including inland marshes (van der Valk 1978). Based on this result, one might conclude that disturbance in detrimental to whitetop meadow productivity. However, the long-term effect (1993 results) indicates that the opposite is the case, as the most disturbed plots accumulated more than twice the biomass of control plots by 1993.
In an earlier study of meadow vegetation at Delta Marsh, Neill (1990a,b) hypothesized that decreases in whitetop biomass in the year following nutrient addition were attributable to litter accumulation. He suggests that litter accumulation has a strong influence on marsh productivity. The importance of litter in lacustrine wetlands was also recognized by van der Valk (1986), who stated that litter accumulation may reduce species richness. In a recent review of the ecological effects of plant litter, Facelli and Pickett (1991) point out that only recently have studies focused on the effects of litter on vegetation community structure and dynamics. In marsh ecosystems, the ecological effects of litter are complex. A litter mat intercepts light, shading seeds (and seedlings) and reducing surface evaporation. Litter also acts as a physical barrier to seedling emergence, and may prevent seed from reaching the soil surface. Through interception of the light and rain, litter affects heat and water transfer between soil and atmosphere.
The litter of the major emergent macrophyte species at Delta Marsh, including whitetop, remains standing for some time. Furthermore, the community dominants accumulate considerable quantities of litter, since all aerial biomass is transformed to litter at end of the growing season and biochemical decomposition is limited by low oxygen levels (Polunin 1984). Decomposition rates may be greater when litter contacts the ground (through, for example, snow-packing or trampling by animals).
In Phragmites communis stands, Haslam (1971) concluded that litter accumulation helps prevent invasion by other species, and as such is an important adaptive strategy. More generally, Grime (1979) hypothesizes that dominance of some plant species in productive habitats is attributable to their ability to accumulate litter. The idea that plant species accumulate litter to deter competitors (through physical and/or chemical means) is only partially supported by this study. The results do, however, suggest that litter accumulation has a strong intraspecific effect; decline in litter accumulation in the most disturbed sites actually resulted in an increase in whitetop biomass. This of course does not negate the possibility that litter accumulation also has a strong interspecific effect. Note, however, that the study by Neill (1990a,b) suggests that excessive litter accumulation by whitetop may actually be detrimental to the species. In Phragmites communis stands, removal of litter increased winter bud mortality, resulting in the release of apical dominance and an increase in density (Haslam 1971). The strong insulative effect of litter may also delay growth in the early spring, however.
This study has demonstrated that a disturbance results in a decrease in whitetop biomass, while at the same time affording the opportunity for ruderal species to germinate, establish and grow. The species invading disturbed patches are primarily opportunistic ruderals that are found in the seed bank (van der Valk and Davis 1978). In lacustrine marshes, such species may 'lie in wait' in the seed bank for considerable period of time (Welling et al. 1988a,b). A 'quiltwork' of disturbance patches in the landscape may be essential to the maintenance of species diversity and richness in lacustrine marshes. However, patches are soon recolonized by the dominant species, usually through clonal growth of roots or rhizomes. It is therefore important that patches are continually formed, and that the species invading such patches are fast-growing, prolific seed producers.
This project could not have been completed without the help and enthusiastic support of Charles Burchill, Krista Copeland, David Ingleson, Geoff Jones, Alex McIlraith and Mike Shaw. The cooperation of the staff of the University Field Station was greatly appreciated. This research was supported by Natural Sciences and Engineering Research Council individual operating grant A-3140.
Facelli, J. M. and Pickett, S. T. A. 1991. Plant litter: its dynamics and effects on plant community structure. Bot. Rev. 57: 1-32.
Grime, J. P. 1979. Plant strategies and vegetative processes. Wiley, New York.
Haslam, S. M. 1971. Community regulation in Phragmites communis Trim. I. Monodominant stands. J. Ecol. 59: 65-73.
Kenkel, N. C. 1993. Ecohydrological changes and vegetation dynamics in the Delta Marsh: preliminary discussion. University Field Station (Delta Marsh), Annual Report. 27: 57-60.
Kvalseth, T. O. 1991. Note on biological diversity, evenness, and homogeneity measures. Oikos 62: 123-127.
Löve, A. and D. Löve. 1954. Vegetation of a prairie marsh. Bull. Torrey Bot. Club 81: 16-34.
Neill, C. 1990a. Effects of nutrients and water levels on emergent macrophyte biomass in a prairie marsh. Can. J. Bot. 68: 1007-1014.
Neill, C. 1990b. Effects of nutrients and water levels on species composition in prairie whitetop (Scolochloa festucacea) marshes. Can. J. Bot. 68: 1015-1020.
Polunin, N. V. C. 1984. The decomposition of emergent macrophytes in fresh water. Adv. Ecol. Res. 14: 115-166.
Smith, A. L. 1973. Life cycle of the marsh grass, Scolochloa festucacea. Can. J. Bot. 51: 1661-1668.
Spence, D. H. 1982. The zonation of plants in fresh water lakes. Adv. Ecol. Res. 12: 37-125.
van der Valk, A. G. 1981. Succession in wetlands: a Gleasonian approach. Ecology 62: 688-696.
van der Valk, A. G. 1986. The impact of litter and annual plants on recruitment from the seed bank of a lacustrine wetland. Aquat. Bot. 24: 13-26.
van der Valk, A. G. and Davis, C. B. 1978. The role of seed banks in the vegetation dynamics of prairie glacial marshes. Ecology 59: 322-335.
Walker, J. M. 1965. Vegetation changes with falling water levels in the Delta Marsh, Manitoba. Ph.D. thesis, University of Manitoba, Winnipeg.
Welling, C. H., Pederson, R. L. and van der Valk, A. G. 1988. Recruitment from the seed bank and the development of zonation of emergent vegetation during a drawdown in a prairie wetland. J. Ecol. 76: 483-496.
Welling, C.H., Pederson, R. L. and van der Valk, A. G. 1988. Temporal patterns in recruitment from the seed bank during drawdowns in a prairie wetland. J. Appl. Ecol. 25: 999-1007.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}