Sharon Gill
Department of Zoology, University of Manitoba
Winnipeg, Manitoba, Canada
E-mail:
|
The function of Yellow Warbler alarm calls
Sharon Gill Department of Zoology, University of Manitoba Winnipeg, Manitoba, Canada E-mail: |
|
Predators pose the greatest threat to the nesting effort of most avian species (e.g., Goossen and Sealy 1982). Approximately 55% and 66% of egg and nestling losses, respectively, are due to predation (Rickelefs 1969). Many passerine nests are also threatened by brood parasites, birds that lay their eggs in the nests of other species, the hosts (Friedmann 1929). The Brown-headed Cowbird (Molothrus ater) is a widespread North American brood parasite (Mayfield 1965), that is known to have laid eggs in the nests of over 200 species (Friedmann and Kiff 1985). Brown-headed Cowbirds usually remove a host egg around the time of parasitism (Sealy 1992). In addition, cowbird nestlings often outcompete host nestlings for food (Friedmann 1963). Whereas predators threaten nests throughout the nesting cycle (Andersson et al. 1980), cowbirds threaten host nests mainly during the hosts' egg-laying and early incubation stages, which is when cowbirds lay most of their eggs (Rothstein 1975).
When defending their nests against predators and brood parasites, nest owners typically utter alarm calls (e.g., Nice 1937; Edwards et al. 1949, 1950; Smith and Hoskings 1955; Seppa 1969; Greig-Smith 1980; East 1981; Knight and Temple 1986a; Hobson and Sealy 1989; Duckworth 1991; Neudorf and Sealy 1992; Mark and Stutchbury 1994). Cowbirds and avian nest predators, such as Common Grackles (Quiscalus quiscula), commonly elicit the same alarm calls from nest owners (e.g., Neudorf and Sealy 1992). Cedar Waxwings (Bombycilla cedrorum) uttered 'disturbance' calls with equal frequency to cowbird and grackle models presented at their nests. In some species, predators may elicit other alarm calls, in addition to those elicited by cowbirds. Gray Catbirds (Dumetella carolinensis) emitted 'mew' calls to both cowbird and grackle models, but primarily gave 'screams' to grackles (Neudorf and Sealy 1992). Few species give unique alarm calls in response to brood parasites (Edwards et al. 1949, 1950; Smith and Hoskings 1955; Seppa 1969; Duckworth 1991; Hobson and Sealy 1989). Most species that do so are hosts of Common Cuckoos (Cuculus canorus).
Among North American birds parasitized by cowbirds, Yellow Warblers (Dendroica petechia) are the only species known to respond to cowbirds with a unique alarm call, the 'seet' call (Hobson and Sealy 1989). This call is given infrequently in response to other intruders such as mammalian nest predators, humans, non-threatening intruders, and conspecific intruders, each of which primarily elicits a structurally and aurally distinct alarm call, the 'chip' call (Ficken and Ficken 1965; Reid and Sealy 1986; Hobson et al. 1988; Hobson and Sealy 1989, 1990). Because the response to an avian nest predator is unknown, however, the seet call may be given to all avian intruders that pose a threat to the nest, rather than being a specific call given to cowbirds.
Alarm calls given in response to a particular stimulus in the environment are called 'referential signals'. Such signals cause nearby conspecifics to react in a way best suited to avoid predation (Smith 1977). Domestic chickens (Gallus gallus) were more likely to look up and run to cover when 'hawk' or aerial alarm calls were played to them than when ground or mammalian alarm calls were given (Evans et al. 1993). Similarly, vervet monkeys (Cercopithicus aethiops) fled into cover and scanned the sky when an aerial alarm call was sounded, whereas they scanned the ground when a ground alarm call was given (Seyfarth et al. 1980a,b). Whether nest-associated alarm calls denote predators or cowbirds has never been examined. Yellow Warblers are ideal candidates for studying referential signals, because the seet call is primarily elicited by cowbirds. Moreover, Yellow Warblers perform nest-protection behaviour, described as a 'rush to sit in the nest', primarily in response to cowbirds (Hobson and Sealy 1989), such that a unique defensive behaviour is expected in response to the seet call (see Smith 1977; Seyfarth et al. 1980a,b).
Alternatively, so-called referential signals may simply reflect the immediacy of response imposed on the prey by the predator (Owings and Hennessey 1984). Thus, a predator elicits a specific alarm call when it is close to the caller, but elicits a different one when it is farther away. Because studies that showed that Yellow Warblers gave the unique seet call to cowbirds presented models only close to the nest, the seet call may indicate response urgency rather than intruder class (see Pereira and Macedonia 1991).
Several hypotheses have been formulated to explain the function of nest-associated alarm calls. To maintain contact and coordinate behaviour, mates often call when they are around their nests (Benson 1939; Slack 1976; Lein 1980; Howes-Jones and Barlow 1988). Thus alarm calls may signal to the mate the presence of an intruder, or more specifically, a cowbird if the seet call is given. Calls may also be directed towards nestlings, causing them to stop begging and crouch in the nest (Greig-Smith 1980; Knight and Temple 1986a). Chip calls are given frequently during the nestling stage, suggesting that they may function in this regard (Hobson et al. 1988). Seet calls are not predicted to warn nestlings, since they may denote cowbirds that threaten the nest primarily at egg-laying stage (but see DuBois 1957; Tate 1966; Beane and Alford 1990 for observations of cowbirds preying on nestlings).
Nest-associated alarm calls also may function to elicit group mobbing by con- and heterospecifics (Curio 1975; Klump and Shalter 1984; Howes-Jones and Barlow 1988; Hobson and Sealy 1989). If Yellow Warbler seet calls function to elicit group mobbing, species which are threatened by cowbirds would be primarily expected to respond (i.e. 'acceptors'). By contrast, if chip calls signal a nest predator, all individuals nesting in an area might be expected to respond.
Finally, alarm calls may function as distraction displays or may act to enhance the effectiveness of distraction displays (Greig-Smith 1980; Knight and Temple 1986a; Montgomerie and Weatherhead 1988). Yellow Warblers give conspicuous distraction displays when models are presented to their nests (Hobson and Sealy 1989). Alarm calls given during such displays may help draw the intruder away from the nest (Montogomerie and Weatherhead 1988).
The objectives of this study were to determine whether: (1) Yellow Warblers utter seet calls only to cowbirds and not to avian nest predators; (2) the seet call is a referential signal denoting cowbirds; (3) Yellow Warbler alarm calls function to warn mates and nestlings of approaching danger; (4) alarm calls function to elicit mobbing by con- and heterospecifics; and (5) alarm calls function as or enhance distraction displays.
At Delta Marsh and Portage Country Club, I located Yellow Warbler nests in the dune-ridge forest that separates Lake Manitoba from the surrounding marsh (MacKenzie 1982). These nests were monitored by G. McMaster or myself every day until clutch completion and every 2-4 days until the young fledged or the nests failed.
I performed three experiments in the 1993 field season. The first aspect of my study consisted of presenting taxidermic mounts of a Fox Sparrow (Passerella iliaca), a Common Grackle (Quiscalus quiscula), and a female Brown-headed Cowbird to nesting Yellow Warblers at the egg-laying and nestling stages. Fox Sparrows migrate through the study and, as such, present a semi-novel stimulus to Yellow Warblers at Delta Marsh. Grackles are the smallest-known avian predator of both eggs and nestlings (Sealy, unpubl. data). At the egg-laying stage, I tested nests from the second day of laying (two eggs) to the third day of incubation. I tested nests with nestlings at least two days old, but no older than seven days old. Yellow Warbler nestlings 8-10 days old will leave the nest prematurely if approached (pers. obs.). Concurrent with model testing, an assistant recorded vocalizations elicited by the model. The assistant was hidden in a blind that was positioned as close to the nest as possible without disturbing the nest owners. In most cases I was also hidden in a blind; otherwise, I positioned myself as far back as possible while maintaining a clear view of the nest. We set up the blinds 15 minutes prior to model testing to allow the nest owners to habituate to them.
After habituation time and if the nest owners were out of the area, I quickly positioned the model approximately 0.5 m from the nest. I clipped models to vegetation and positioned them facing the nest. If the nest owners were in the area after the 15-minute period, I waited to present the model until both left the area. I presented the models in a random order and separated each test by at least 15 minutes to reduce habituation or carry-over aggression (Knight and Temple 1986b). Once the nest owner responded, I started behavioural observations, while my assistant began recording any vocalizations elicited. For five minutes, I recorded the nest owners' responses on audio tapes using a hand-held tape-recorder and transcribed the tapes later. I recorded all responses displayed by both male and female nest owners, but in the present report I focus on alarm calls and nest-protection behaviour. I recorded nest-protection behaviour as the number of 10-second intervals in which it occurred, and alarm calls as the number of times they were given in the 5-minute trial. If both nest owners called during the model trial I could not assign vocalizations to one sex. Therefore, I combined male and female calling, and compiled the proportion of calling by each sex at both egg-laying and nestling stages.
Vocalizations elicited by the models were recorded by my assistant using a Uher 4000 Report-L, Sennheiser ME 88 microphone with fixed windscreen and K3 low frequency filter, and Ampex Precision Magnetic tapes. The Sennheiser microphone is highly directional, and suitable when the microphone cannot be positioned close to the sound source, as was the case during model trials. Tape speed was always set at 19 cm / sec to make the best quality recording possible (Spector 1991). The filter suppresses low and high levels of noise at positions II and III, respectively. I set the filter at position II during low wind conditions, and at position III when it was windy. I recorded the filter position used in all cases.
In the second set of model trials I presented female cowbird models at three distances from the nest: 0.5, 2.5 and 4.5 m (n=22). All protocols were as above except that vocalizations were not recorded, models were occasionally positioned on poles, and nests were tested only during egg-laying stage. If poles were used I positioned them 15 minutes prior to the first trial and repositioned them at the end of each trial. If nest owners did not respond within 30 minutes, I removed the model and recorded this as no response.
The second aspect of my study consisted of playing back seet and chip calls to nest owners. Playbacks consisted of 1 minute of seet or chip calling. This length of playback is appropriate to prevent habituation (Falls 1982). Moreover, parasitism by cowbirds (Sealy et al., in press), and predation (pers. obs.), occur within 1-2 mins. From the recordings made during model testing, I selected a section of calling 2-30 seconds long from which I made a one minute playback using the Uher Report-L recorder, a Sony TCM-5000EV recorder and Sony Metal SR tapes. The template varied considerably in length because nest owners varied in the length of time they spent calling. I also recorded one minute of background noise that was used as a control.
I performed the playback experiment one or two days after model testing. Accordingly, both egg-laying and nestling stages were tested. Playbacks were delayed by two days only when it rained. Sample sizes here were smaller than in model presentation because some tested nests were depredated over night (see results). Fifteen minutes prior to testing I set out a blind and placed an Audio-Technica amplified speaker 1 m below the nest, either on the ground or in a crotch of a tree. The speaker positions are appropriate as Yellow Warblers vocalized from the ground and from perches (pers. obs.). The speaker was connected by a 10-m cord to the Sony TCM-5000EV recorder. I randomized the order of the playbacks. Some nest owners did not utter seet or chip calls during model testing. Therefore, the sample sizes for self call playbacks vary among treatments (seet: egg-laying [n=22], nestling [n=9]; chip: egg-laying [n=18], nestling [n=19]).
One minute before playback, I began behavioural observations. I recorded behaviours directly into a field notebook in which I had delineated seven minutes into 10 second intervals. I recorded male and female behaviours using the same categorization as in model testing, and again focus on the vocalizations emitted and nest-protection behaviour. I started playbacks after one minute of observations except when the female was on the nest. If the female was on the nest, I waited until she left. For five minutes after playback, I recorded the behaviour of the nest owners. Subsequent test calls were played back after five minutes, for a total of ten minutes between successive trials.
In addition to recording the nest owners' behaviours, I attempted to determine nestling response to the playbacks. This proved more difficult than I had anticipated. Yellow Warbler nestlings that are a few days old are difficult to see in the nest. Furthermore, they do not start vocalizing until they are 7-8 days old (pers. obs.). However, nestlings at this age may also start leaving the nest when the nest is checked. Also, the position of most nests did not allow me to see the nestlings. Thus, I provide only anecdotal observations of nestling responses.
Because the data were not normally distributed, I used non-parametric tests to analyze the results. To determine whether model type (sparrow, grackle or cowbird) or distance (0.5 m, 2.5 m or 4.5 m from the nest) influenced the nest defense responses elicited, I ranked the data within each nest (blocks), and then performed a Friedman two-way ANOVA. When significant differences resulted (p < 0.05), I used Fisher's protected least significant difference (FPLSD) test on the ranks to determine which model(s) elicited a significantly different response(s). Because the hypotheses concerning playbacks specify the appropriate comparisons, I used x2 test of homogeneity to determine whether the proportion of females performing each behaviour was influenced by call type.
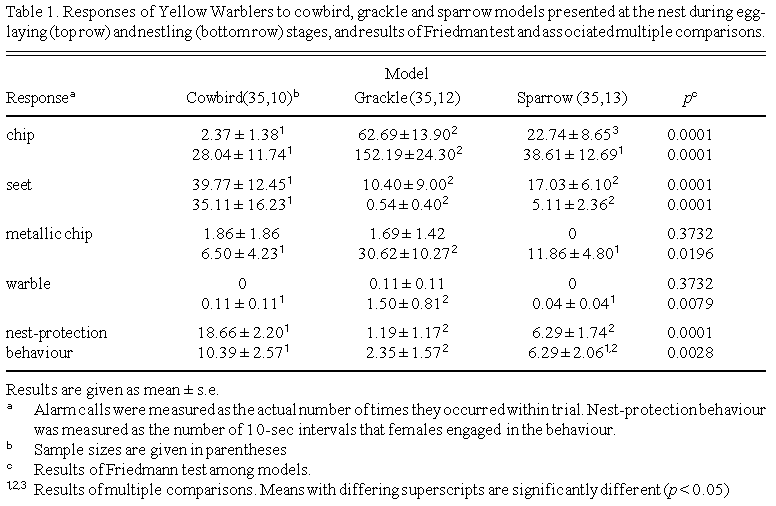
Yellow Warblers frequently vocalized during model presentation. Yellow Warblers uttered significantly more chip calls in response to the grackle model than the other models, but more to the sparrow than the cowbird (Table 1). Conversely, Yellow Warblers gave significantly more seet calls to the cowbird model than either the sparrow or the grackle. Most females uttered seet calls as they rushed to sit in the nest (i.e. nest-protection behaviour). Once females entered their nests they usually remained silent. Yellow Warblers gave also metallic chip calls (Ficken and Ficken 1965) to cowbird and grackle models only, although this was not significant. A previously undescribed vocalization, which I describe as a soft-pitched warble call, was elicited only by the grackle at the egg-laying stage. Warble calls were given rarely and only when the warbler was distraction displaying.
At the nestling stage, Yellow Warblers gave significantly more chip calls to the grackle than to the other models. They uttered more seet calls in response to the cowbird model than the grackle or sparrow models. Both metallic chips and warbles were elicited more by the grackle model than by the cowbird or sparrow. Nest-protection behaviour was performed more frequently by females during cowbird trials than grackle trials.
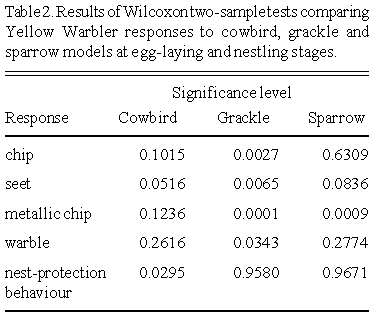
All alarm calls except the seet call increased over the nesting cycle (Table 2). The number of seet calls given towards the cowbird and sparrow models decreased significantly over the nesting cycle. Yellow Warblers gave significantly more chip, metallic chip, and warble calls to the grackle at the nestling stage than at egg-laying stage. Although rarely given towards sparrow model, the number of pink calls elicited by this model increased significantly from egg-laying to nestling stages. Time spent in the nest by females in response to the cowbird model decreased significantly over the breeding cycle.
Warblers uttered chip calls more frequently when the model was farther from the nest, but this was not significant (Table 3). Seet calls were uttered at an equal rate during all trials, given primarily when the model was first sighted. Females sat in the nest more in response to the model placed closest to the nest than at the greater distances.
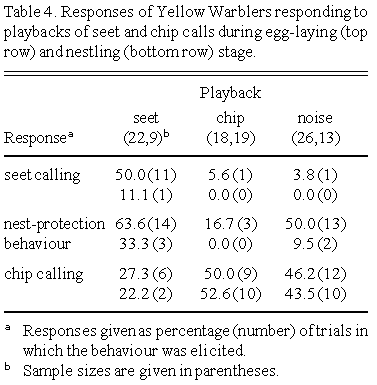
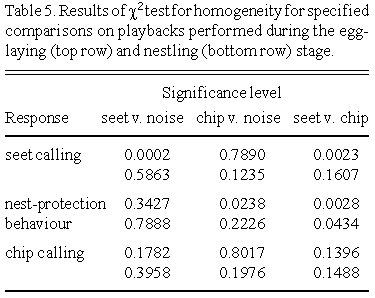
Yellow Warblers uttered significantly more seet calls in response to the seet playback than either the chip or noise playback (Table 4 and Table 5). Although Yellow Warblers sat in the nest significantly more in response to the seet playback than the chip, the proportion of birds sitting in the nest did not differ between the seet and noise playback. Accordingly, females sat in the nest more in response to the noise playback than the chip playback. The proportion of birds uttering chip calls did not differ between the playbacks. At the nestling stage, Yellow Warblers responded similarly to all playbacks.
The proportion of individuals giving seet calls and the proportion of females sitting in the nest in response to the seet call was significantly greater at the egg-laying stage than at the nestling stage (Table 6).
Yellow Warblers responded with four alarm calls to the three models, but each model elicited a different set of vocalizations. The cowbird model elicited primarily seet calls, particularly at the egg-laying stage. Hobson and Sealy (1989) also found that Yellow Warblers gave more seet calls to cowbirds at egg-laying than the other nesting stages. Chip, metallic chip (Ficken and Ficken 1965) and warble calls were infrequently given to the cowbird model at egg laying, but each call was given more frequently over the nesting cycle.
By contrast, the grackle and sparrow models elicited chip calls most frequently, but the number of chip calls given to the grackle model far exceeded those given to the sparrow. Yellow Warblers uttered significantly more chip calls to the grackle at the nesting stage. Metallic chip calls were rarely given at egg-laying stage in response to any model, but increased significantly over the nesting cycle in response to grackle and sparrow models. I also recorded 'warble' calls, which have not been described in the literature. These soft-pitched calls are given rarely at either nesting stages. Warble calls were given most to the grackle model at the nestling stage, and only when the nest owners were distraction displaying.
The vocal responses to the models indicate that Yellow Warblers discriminate between the cowbird and grackle, and that they recognize the unique threats each poses. Yellow Warblers gave the seet call primarily to the cowbird model, and rarely to the grackle model, indicating that the seet call is a specific response to cowbirds rather than uttered in response to any avian intruder threatening the nest. Furthermore, female warblers performed nest-protection most frequently to the cowbird model. These results corroborate previous studies that suggested unique recognition of cowbirds by Yellow Warblers (Robertson and Norman 1977; Folkers 1982; Folkers and Lowther 1985; Burgham and Picman 1989; Hobson and Sealy 1989). Few other studies have examined whether hosts respond differently to nest predators and brood parasites. Neudorf and Sealy (1992) showed that Red-winged Blackbirds discriminate between these distinct nest threats, but that three rejecter species showed little or no discrimination. Duckworth (1991) found that Reed Warblers (Acrocephalus scirpaceus) discriminated between nest predators (Jays Garrulus glandularis) and Common Cuckoos.
Results from the model experiment provide preliminary support that the seet call is a referential signal denoting cowbirds, because the call was given primarily to cowbirds at the egg-laying stage. Furthermore, Yellow Warblers gave the same number of seet calls to the cowbird model at each of the distances it was from the nest, whereas nest-protection behaviour decreased with increasing distance. Thus, seet calls did not vary depending on whether the situation was 'high urgency' (close to the nest) or 'low urgency' (far from the nest; see Pereira and Macedonia 1991). The other alarm calls of Yellow Warblers may reflect response urgency, however. Metallic chip calls are heard mostly in the latter half of model trials, perhaps as the bird escalates the interaction because it has been unable to move the intruder (model) away from the nest. Ficken and Ficken (1965) indicated that chip and metallic chip calls may grade into each other. Similarly, the warble call may reflect an escalation in the interaction.
Calls that are referential signals are predicted to elicit defensive behaviours normally given in an actual encounter with the stimulus (Smith 1977; Seyfarth et al. 1980a,b). Yellow Warblers performed nest-protection behaviour and seet calling when the seet call was played to them. Thus Yellow Warbler seet calls are referential signals denoting cowbirds. This is the first study to show the existence of a referential signal in response to a brood parasite or even a nest threat.
The playback of background noise caused females to sit in the nest in 50% of trials. Because females usually uttered seet calls in association with nest-protection behaviour, I suspect that the responses of females to the noise and seet playbacks are not the same in a functional sense. Females rushed back to the nest seet calling in response to the seet playback. By contrast, females appeared to be returning to the nest to incubate or brood during noise playbacks, and never uttered seet calls during this return. Therefore seet and noise playbacks elicited two very different behaviours.
Chip call playback did not elicit intense responses by Yellow Warblers. However, females did not return to the nest during chip call playback suggesting that it might have a 'stay away' function. This was more apparent at the nestling stage, when few females approached the speaker at all. During testing with the grackle model, nest owners usually stayed over two meters from the model. Perhaps the chip calls communicate the presence of a predator, such that nest owners do not approach too closely. Cheney and Seyfarth (1982) showed that vervet monkey responses to 'grunt' calls are subtle, consisting mostly of movement to and away from the speaker.
I tested three hypotheses on the function of alarm calling in Yellow Warblers: (1) Alarm calls elicit group mobbing; (2) Alarm calls warn mate and offspring of potential danger; and (3) Alarm calls act as a distraction display. I will go over each hypothesis separately and present evidence gathered from my experiments.
Hobson and Sealy (1989) proposed that Yellow Warbler seet and chip calls may cause group mobbing, but conceded that they never recorded mobbing during model trials. During the playback experiment, I never recorded birds other than the nest owner(s) responding. By contrast, during 34 model trials, I frequently recorded individuals other than the nest owners responding. Most of the birds that responded were Yellow Warblers, but Gray Catbirds, Red-winged Blackbirds (Ageliaus phoeniceus), and Song Sparrows (Melospiza melodia) also were drawn to the area. Calling during the grackle model presentation drew in birds most frequently (21 / 34 cases). Furthermore, a Common Grackle I observed depredating a Yellow Warbler nest attracted four other warblers, plus a Northern Oriole (Icterus galbula) and Gray Catbird (pers. obs.).
Birds other than nest owners may require additional visual cues to identify the stimulus before they respond. Although the seet call will identify to Yellow Warblers that the intruder is a cowbird, other warblers did not respond to the playback and only rarely appeared during presentation of the cowbird model (2 / 34 cases). Host aggression may lead cowbirds to nests (see Robertson and Norman 1976,1977; Smith et al. 1984; Gill et al., unpubl. data), therefore other Yellow Warblers may not risk responding unless their nest is threatened directly.
Chip calls are given in a wide variety of situations, and to various intruders at the nest, both threatening and benign (Ficken and Ficken 1965; Reid and Sealy 1986; Hobson et al. 1988; Hobson and Sealy 1989, 1990). Because they are given in so many contexts, warblers may rely on contextual information to determine the meaning of the call (Smith 1977; Lein 1980). In the absence of such information, warblers, including nest owners, may not respond. Alternatively, the structure of chip calls may vary with context, such that contextual information is encoded in the call (Stone and Trost 1991). If this occurs then chip calls given to the grackle would elicit different responses than those given when foraging or leaving the nest. Whether Yellow Warblers respond differently to chip calls recorded from different contexts needs to be determined.
The second hypothesis on the function of Yellow Warbler alarm calls that I tested considered whether the calls warn the mate and offspring of an intruder in the area. I gathered only anecdotal and equivocal observations on whether the alarm calls warn nestlings of approaching danger. Some nestlings retracted their heads during chip call playbacks, but other nestlings did not. I can conclude nothing from these meager observations. However, I have suggestive evidence that the chip call may function to warn nestlings. Chip calling without apparent stimuli increases in frequency over the nesting cycle (pers. obs.). Additionally, the farther the female was away from her nest, the more chip calls she gave (pers. obs.). This suggests that calling may indicate to the nestlings that the female is not near the nest. The increased frequency of chip calling in response to the grackle model over the nesting cycle may reflect communication to the nestlings as well (see Gyger et al. 1986). Black-billed magpies (Pica pica) increased their alarm calling as their nestlings grew older and louder, presumably to keep them quiet (Buitron 1983). The seet call is not expected to communicate anything to nestlings, as cowbirds present little threat to nests with nestlings.
Alarm calls may also warn the mate of impending danger. Both seet and chip call playback attracted female and male nest owners back to their nest. Seet calls also elicited nest-protection behaviour, another response given primarily to cowbirds. Thus these alarm calls may be used by nest owners, especially females, to bring their mate back to the nest, and to solicit aid in nest defense from the mate. Small passerines may defend the nest most successfully when two parents are present (Moksnes et al. 1990).
The third hypothesis for alarm call function is that calls act as a distraction display, drawing the intruder away from the nest (Greig-Smith 1980; Knight and Temple 1986a), or enhancing the effectiveness of distraction displays (Montgomerie and Weatherhead 1988). Although not tested directly, this is likely to be important for Yellow Warblers. During egg-laying stage, most warblers were silent when they displayed. Nest owners stopped calling before they engaged in displaying, calling only after they stopped displaying. Similarly, warblers distraction displaying during the nestling stage were initially silent, primarily vocalizing only after displaying was unsuccessful. Yellow Warblers unable to draw the intruder (model) away, started alarm calling later in the trial. This suggests that the nest owners were attempting to increase effectiveness of distraction display by drawing more attention to themselves by calling (Montgomerie and Weatherhead 1988). The soft-pitched warble call may be especially effective in this regard. This call sounds like it is coming from an injured animal. That it was given most in response to the grackle model during the nestling stage, and only while the nest owner was distraction displaying, supports this contention. The grackle represents a greater threat to the nest than either the sparrow or cowbird model at this time (Patterson et al. 1980).
I thank Dr. Spencer G. Sealy for guidance and assistance throughout the summer and beyond. I appreciate the care taken by D. Beattie, K. Caldwell, D. Froese and L. Zdrill in recording the alarm calls. G. McMaster was invaluable to me as he found and monitored most of the nests I used in my study. P. Grieef and G. McMaster discussed aspects of my project with me, for which I am grateful. A big thank you to the staff at Delta Marsh for their hospitality. The officers of the Portage Country Club kindly permitted me to do portions of my work on their property. Statistical advice was provided by Llwellyn Armstrong of the University of Manitoba Statistical Advisory Service. Funding was provided by an NSERC Research Grant to Dr. S.G. Sealy and an NSERC Postgraduate Scholarship to myself.
Andersson, M., Wiklund, C. G. and Rundgen, H. 1980. Parental defence of offspring: a model and an example. Anim. Behav. 28: 536-542.
Beane, J. C. and Alford, S. L. 1990. Destruction of a Pine Warbler brood by an adult cowbird. Chat 54:85-87.
Benson, M. H. 1939. A study of the American Redstart (Setophaga ruticilla Swainson). MSc thesis, Cornell University, Ithaca, NY.
Buitron, D. 1983. Variability in the responses of Black-billed Magpies to natural predators. Behaviour 87: 209-236.
Burgham, M. C. J. and Picman, J. 1989. Effect of Brown-headed Cowbirds on the evolution of Yellow Warbler anti-parasite strategies. Anim. Behav. 38: 298-308
Cheney, D. L. and Seyfarth, R. M. 1982. How vervet monkeys perceive their grunts: field playback experiments. Anim. Behav. 30: 739-751.
Curio, E. 1975. The functional organization of anti-predator behaviour in the Pied Flycatcher: a study of avian visual perception. Anim. Behav. 23: 1-115.
DuBois, A. D. 1956. A cowbird incident. Auk 73: 286.
Duckworth, J. W. 1991. Responses of breeding Reed Warblers Acrocephalus scirpaceus to mounts of Sparrowhawk Accipiter nisus, Cuckoo Cuculus canorus and Jay Garrulus glandarius. Ibis 133: 68-74.
Edwards, G., Hoskings, E. and Smith, S. 1949. Reactions of some passerine birds to a stuffed cuckoo. Brit. Birds 42: 13-19.
Edwards, G., Hoskings, E. and Smith, S. 1950. Reactions of some passerine birds to a stuffed cuckoo. II. A detailed study of the Willow Warbler. Brit. Birds 43: 144-150.
Evans, C. S., Evans, L. and Marler, P. 1993. On the meaning of alarm calls: functional reference in an avian vocal system. Anim. Behav. 46: 23-38.
Falls, J. B. 1982. Individual recognition by sound in birds. In Kroodsma, D. E. and Miller, E. H. (eds.), Acoustic Communication in Birds, Vol. 2: 237-278. New York: Academic Press.
Ficken, M. S. and Ficken, R. W. 1965. Comparative ethology of the Chestnut-sided Warbler, Yellow Warbler, and American Redstart. Wilson Bull. 77: 363-375.
Folkers, K. L. and Lowther, P. E. 1985. Responses of nesting Red-winged Blackbirds and Yellow Warblers to Brown-headed Cowbirds. J. Field Ornithol. 56: 175-175.
Folkers, K. L. 1982. Host behavioral defenses to cowbird parasitism. Bull. Kansas Ornithol. Soc. 33: 32-34.
Friedmann, H. 1929. The Cowbirds. A Study in the Biology of Social Parasitism. Springfield: Charles C. Thomas.
Friedmann, H. 1963. Host relations of the parasitic cowbirds. US Natl. Mus. Bull. 223.
Friedmann, H. and Kiff, L. F. 1985. The parasitic cowbirds and their hosts. Proc. West. Foundn. Vert. Zool. 2: 226-302.
Goossen, J. P. and Sealy, S. G. 1982. Production of young in a dense nesting population of Yellow Warblers, Dendroica petechia, in Manitoba. Can. Field-Nat. 96: 189-199.
Greig-Smith, P. W. 1980. Parental investment in nest defence by Stonechats (Saxicola torquata). Anim. Behav. 28: 604-619.
Gyger, M., Karakashian, S. J. and Marler, P. 1986. Avian alarm calling: is there an audience effect? Anim. Behav. 34: 1570-1572.
Hobson, K. A., Bouchart, M. L. and Sealy, S. G. 1988. Responses of naive Yellow Warblers to a novel nest predator. Anim. Behav. 36: 1823-1830.
Hobson, K. A. and Sealy, S. G. 1989. Responses of Yellow Warblers to the threat of cowbird parasitism. Anim. Behav. 38: 510-519.
Hobson, K. A. and Sealy, S. G. 1989. Female-female aggression in polygynous nesting Yellow Warblers. Wilson Bull. 101: 84-86.
Hobson, K. A. and Sealy, S. G. 1990. Breeding-season aggression of female Yellow Warblers to models of male and female conspecific intruders. Anim. Behav. 39: 809-811.
Howes-Jones, D. and Barlow, J. 1988. The structure of the call note system of the Warbling Vireo. Royal Ontario Museum Life Sciences Contribution 151: iv + 36 pp.
Klump, G. M. and Shalter, M. D. 1984. Acoustic behaviour of birds and mammals in the predator context. Z. Tierpsychol. 66: 189-226.
Knight, R. L. and Temple, S. A. 1986a. Nest defence in the American Goldfinch. Anim. Behav. 34: 887-897.
Knight, R. L. and Temple, S. A. 1986b. Methodological problems in studies of avian nest defence. Anim. Behav. 34: 561-566.
Lein, M. R. 1980. Display behavior of Ovenbirds (Seiurus aurocapillus) I. Non-song vocalizations. Wilson Bull. 92: 312-329.
MacKenzie, D. I. 1982. The dune-ridge forest, Delta Marsh, Manitoba: overstory vegetation and soil patterns. Can. Field-Nat. 96: 61-68.
Mark, D. and Stutchbury, B. J. 1994. Response of a forest-interior songbird to the threat of cowbird parasitism. Anim. behav. 47: 275-280.
Mayfield, H. 1965. The Brown-headed Cowbird with old and new hosts. Living Bird 4: 13-28.
Moksnes, A., Roskaft, E., Braa, A. T., Korsnes, L., Lampe, H. M. and Pederson, H. C. 1990. Behavioural responses of potenetial hosts towards artificial cuckoo eggs and dummies. Behaviour 116: 64-89.
Montgomerie, R. D. and Weatherhead, P. J. 1988. Risks and rewards of nest defence by parent birds. Quart. Rev. Biol. 63: 167-187.
Neudorf, D. L. and Sealy, S. G. 1992. Reactions of four passerine species to threats of predation and cowbird parasitis: enemy recognition or generalized responses? Behaviour 123: 84-105.
Nice, M. M. 1937. Studies of the Life History of the Song Sparrow. Vol. 2. Trans. Linn. Soc. NY. 4: 246 pp.
Owings, D. H. and Hennessy, D. F. 1984. The importance of variation in sciurid visual and vocal communication. In Murie, J. O. and Mitchener, G. R. (eds.) The Biology of Ground-dwelling Squirrels: 169-200. Univ. Nebraska Press.
Patterson, T. L., L. Petrinovich and James, D. K. 1980. Reproductive value and appropriateness of responses to predators by White-crowned Sparrows. Behav. Ecol. Sociobiol. 7: 227-231.
Pereira, M. E. and Macedonia, J. M. 1991. Ringtailed lemur anti-predator calls denote predator class, not response urgency. Anim. Behav. 41: 543-544.
Reid, M. L. and Sealy, S. G. 1986. Behavior of a polygynous yearling Yellow Warbler. Wilson Bull. 98: 315-317.
Rickelefs, R. E. 1969. An analysis of nesting mortality in birds. Smithsonian Contr. Zool. no. 9.
Robertson, R. J. and Norman, R. F. 1976. Behavioral defenses to brood parasitism by potential hosts of the Brown-headed Cowbird. Condor 78: 166-173.
Robertson, R. J. and Norman, R. F. 1977. The function and evolution of aggressive host behavior towards the Brown-headed Cowbird (Molothrus ater). Can. J. Zool. 55: 508-518.
Rothstein, S. I. 1975. An experimental and teleonomic investigation of avian brood parasitism. Condor 77: 250-271.
Sealy, S. G. 1992. Removal of Yellow Warbler eggs in association with cowbird parasitism. Condor 94: 40-54.
Seppa, V. 1969. The cuckoo's ability to find a nest where it can lay an egg. Ornis Fenn. 46: 78-80.
Seyfarth, R. M., Cheney, D. L. and Marler, P. 1980a. Monkey responses to three different alarm calls: evidence of predator classification and semantic communication. Science 210: 801-803.
Seyfarth, R. M., Cheney, D. L. and Marler, P. 1980b. Vervet monkey alarm calls: semantic communication in a free-ranging primate. Anim. Behav. 28: 1070-1094.
Slack, R. D. 1976. Nest guarding behavior by male Gray Catbirds. Auk 93: 292-300.
Smith, J. N. M., Arcese, P. and McLean, I. G. 1984. Age, experience and enemy recognition in wild Song Sparrows. Behav. Ecol. Sociobiol. 14: 101-106.
Smith, S. and Hosking, E. 1955. Birds Fighting. Experimental Studies of Aggressive Displays of Some Birds. London: Faber and Faber.
Smith, W. J. 1977. The Behavior of Communicating. Harvard Univ. Press.
Spector, D. A. 1991. The singing behaviour of Yellow Warblers. Behaviour 117: 27-32.
Stone, E. and Trost, C. H. 1991. Predators, risks and context for mobbing and alarm call in Black-billed Magpies. Anim. Behav. 41: 633-638.
Tate, J., Jr. 1967. Cowbird removes warbler nestling from nest. Auk 84: 422.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}