L. Gordon Goldsborough
Department of Botany, Brandon University
Brandon, Manitoba, Canada
| Effects of hexazinone on water
chemistry and periphyton biomass and productivity in large littoral enclosures
L. Gordon Goldsborough Department of Botany, Brandon University Brandon, Manitoba, Canada |
|
Periphyton communities develop through successional processes analogous to those of macroscopic plants in much shorter periods of time. Therefore, manipulations of lentic periphyton may provide useful models for the responses of plant communities generally to anthropogenic stress. The increasing use of herbicides in agriculture and forestry is resulting in widespread contamination of standing waters (e.g., Muir & Grift 1987). Given that plants are the primary targets of these compounds and that eutrophic aquatic systems are hypothesized to be producer limited (McQueen et al. 1986, Power 1992), the extent to which aquatic residues will impact on long term ecosystem production is an important concern that merits attention.
I conducted manipulative experiments in the Blind Channel using replicated littoral enclosures to investigate the herbicide-induced stress responses of periphyton. The herbicide chosen for these experiments was hexazinone (3-cyclohexyl-6-dimethylamino-1-methyl-1,3,5-triazine-2,4(1H,3H)-dione), a triazine-class herbicide whose mechanism of action is blockage of electron flow in photosystem II of photosynthesis. This chemical was chosen for three reasons: it is highly water soluble (33 g·Lą; WSSA 1983) and essentially nonvolatile, so it should have a long residence time in the water column, thereby minimizing effects of changing concentration on observed phytotoxicity. Its well-defined mechanism of action means that it should have demonstrable primary effects on photosynthetic activity of a periphyton community, although the degree to which individual species in the community respond may vary; triazine-tolerant agricultural weeds have been reported. Finally, hexazinone has the potential to contaminate lentic waters, since it is used as a forestry herbicide in Canada for site preparation and conifer release in plantations (Campbell 1990), and has been shown to occur in surface and groundwater after operational use (Williamson 1988). It has also been tested for efficacy in controlling aquatic weeds in the USA (Anderson 1981). I tested a hypothesis that a stressor that inhibits photosynthesis will reduce the rate of carbon fixation and biomass of a natural periphyton community. I also tested the hypothesis that, with prolonged exposure to the stressor, effects on periphyton biomass and productivity will diminish or even reverse through increased community resistance, similar to that reported by Blanck and Wängberg (1988). "Resistance" is used here to refer to decreased response by a multispecific assemblage, whereas "tolerance" refers to unispecific responses.
Four square enclosures, 5 m on a side, were deployed in a square in the Blind Channel in early June, 1991. The perimeter of each enclosure (designated E1 to E4) consisted of four 5.3 m long x 30 cm wide wooden platforms supported by rectangular Styrofoam blocks, which held the resulting square frame about 20 cm above the water surface. The platforms provided a walkway, and a moveable wooden catwalk was positioned between opposite sides of the frame to facilitate sample collection from any position within the enclosure. A square curtain (5 m x 5 m x 180 cm high) made of 6-mil woven polyethylene was attached to the inside surface of the floating frame. The lower edge of the curtain was folded to make a pocket around the entire circumference, in which iron bars were placed. As the curtain was lowered in the water column, the weight of the bars caused the curtain to penetrate the sediments about 20 cm, thereby isolating the water within the enclosure from the surrounding channel. Water depth in the enclosures on the date of curtain deployment (13 June) was 80 cm, yielding an enclosed water volume of 20 mł with a wall surface area to volume ratio of about 0.8 m˛·mł. Since the height of the flexible curtain was greater than the maximum depth of water at the site, the enclosed water columns remained isolated throughout the experiment, although limited water exchange could occur through the sediments. An effort was made to remove fish from the enclosures using Gee minnow traps in the four weeks after curtain deployment. No attempt was made to remove macrophytes from the enclosures.
Previous experiments with enclosures at the study site have shown that differences in enclosed water chemistry and plant productivity develop in replicated enclosures, despite concerted efforts to ensure that all are treated similarly (Goldsborough, in prep.). To characterize fully the pertinent inter-enclosure differences that might interfere with differentiation of treatment effects from background variability, treatment of enclosures with herbicide was delayed for 34 days after deployment of the enclosure curtains. During this time, water and periphyton samples were collected regularly (see below). Two of the four enclosures were randomly selected to receive the same concentration of hexazinone, while the remaining two enclosures served as untreated references.
The hexazinone concentration to be added to treatment enclosures was chosen on the basis of static toxicity experiments (Goldsborough and Forster, submitted). A quantity of Velpar L(r) sufficient to achieve this concentration in the estimated enclosure volume was diluted in 1L filtered marsh water and added slowly to the surface of each treatment enclosure on 24 July 1991. The water in each enclosure was mixed immediately after treatment by drawing a wide plastic paddle across the water surface several times. The date of treatment was designated as day 0, and sampling of water and periphyton from all enclosures continued for the next 37 days. The experiment was terminated on 31 August 1991.
Surface water samples (10-15 cm depth) were collected from each enclosure for ten weeks at 3-4 day intervals from day -30 (30 days before treatment) to day 37. These samples were analyzed chemically for specific conductivity, pH, total titratable alkalinity, ammonia-N, nitrate-N, soluble reactive phosphorus (SRP), and dissolved silica using methods of Stainton et al. (1977) and APHA (1980). Additional samples were filtered through neutralized Whatman GF/C filters and retained for total planktonic chlorophyll a analysis after the method of Marker et al. (1980), using 90% methanol as a pigment extractant. Water temperature and dissolved oxygen levels at 20 cm depth were measured using a YSI Model 51B meter. Periodic measurements at 40 cm and 60 cm depth showed no consistent evidence of depth profiles of either temperature or oxygen. PAR extinction profiles in the center of each enclosure were measured using a Li-Cor LI-1000 datalogger receiving simultaneous input from a LI-185S underwater sensor and a LI-190 terrestrial sensor to correct for temporal variability in cloud cover.
Four hours after herbicide treatment, and at weekly intervals to day 35, 1L whole water samples were collected for herbicide residue analysis in rinsed amber glass bottles at two sites in each treated enclosure, and from one site in each untreated enclosure. The samples were shipped on ice within three hours of collection to Xenos Laboratories Inc. (Ottawa, Ontario), where they were frozen until analysis. The HPLC procedure for analysis of hexazinone was modified from Reupert et al. (1990), in which the samples were brought to room temperature, shaken and a 250µL aliquot of a 5 mL filtered (0.45 µm pore size) sample was injected directly onto the column. The HPLC system consisted of a Varian 9010 pump, a Varian 9050 UV detector (operated at 254 nm) and a Star HPLC Data system. The column was 25 cm x 4.6 mm, 10µ Partisil ODS2 (Whatman), and the mobile phase was 50:50 acetonitrile:ammonium acetate (0.01M) at a flow rate of 1 mL·miną. The retention time for hexazinone was 7.6 min and the limit of detection was 25 µg·Lą. Recovery of a series of fortified control samples was 96±13% (n=9). Metabolites of hexazinone could not be analyzed due to absence of standards, but consistent HPLC peaks other than those of the parent compound were not observed.
Artificial substrata similar to those used in preliminary toxicity experiments (described above) were positioned vertically (30 cm in the sediments) in each enclosure on 20 June (day -34). Prior to placement, each 90 cm long substratum was notched with a small saw to facilitate the collection of discrete sub-segments. Two 2.5 cm long segments at a position on each substratum corresponding to 15-20 cm above the sediment interface, and two 2.5 cm segments at 45-50 cm, were intended for algal carbon fixation measurements. Two 5 cm segments above and below segments for carbon fixation rate measurements were pooled for a single chlorophyll analysis, which served as an indicator of periphyton biomass. The purpose of sampling at two positions on each substratum was to investigate vertical profiles of periphyton productivity and biomass. All substrata were cleaned with 100% methanol prior to placement to remove contaminants arising from hand contact. Seventy-eight substrata were positioned in each enclosure in a regular 10 x 10 regular lattice pattern. Substratum sampling commenced after 7 days of colonization (day -27) and continued at weekly intervals to day 36. At each sampling interval, six substrata were removed randomly from each enclosure by sampling without replacement from a pool of random numbers between 1 and 100.
Segments sampled for carbon fixation rate measurements were placed into glass culture tubes containing 25 mL filtered (1X GF/C) water from the enclosure from which the substrata were sampled that was collected an hour prior to substratum sampling. Two segments sampled at one depth on each substratum were placed into paired clear and blackened tubes. Segments sampled for chlorophyll analyses were placed into empty glass tubes. All tubes were transported to the laboratory, where tubes for chlorophyll analysis were frozen. Tubes for carbon fixation measurement were inoculated with 0.5 NaH14CO3 (37 kBq·mLą) and incubated for 3-4 hours at 25±1°C at 500-600 µE·m-˛·h-ą. Determination of sample radioactivity and calculation of carbon fixation rates (µgC·cm˛·hą) followed the methods of Goldsborough and Robinson (1983). Samples collected for carbon fixation rate determination on day -20 were lost due to defective radiolabel.
Chlorophyll analysis was performed by adding 10 mL 90% neutralized methanol to thawed substratum segments, eluting pigments overnight at room temperature in the dark, and measuring absorbance of the extract at 665 nm and 750 nm, before and after acidification with 10-3N HCl, using a Milton-Roy Spectronic 601 spectrophotometer. Concentrations of chlorophyll a and pheophytin a were calculated using the formulae of Marker et al. (1980) and the two values were summed for total chlorophyll, expressed on an areal basis (µg·cm˛). Photosynthetic efficiency was calculated as the ratio of carbon fixation rate to total chlorophyll concentration (µgC·µgChlą·hą), using chlorophyll values for substratum segments nearest to corresponding segments used for carbon fixation rate measurement.
Levels of periphyton biomass and productivity were tested using two-factor analysis of variance (SAS 1986). To guard against the possibility of Type II errors arising from potential temporal autocorrelation between successive samples from a single enclosure, a stricter critical p-value of 0.01 was adopted in significance testing rather than 0.05. Comparison of carbon fixation rates and chlorophyll concentrations of reciprocally exchanged substrata was performed using two-tailed t-tests using a critical p-value of 0.01.
In a four-hour incubation, hexazinone concentrations < 10 µg·Lą had no effect on carbon fixation rate of an intact periphyton community developed for three weeks at the study site. At increasing concentrations, carbon fixation rate was reduced proportionately, to 10% of control samples at 1200 µg·Lą and 7% of controls at 19,200 µg·Lą. The EC50 concentration was estimated as 40 µg·Lą hexazinone. Based on these data, a hexazinone concentration of 200 µg·Lą was chosen for enclosure treatment. Two considerations influenced this choice. From an ecological standpoint, this concentration was predicted, based on its position near the upper asymptote of the dose-response curve, to eliminate from the community all but the most herbicide-tolerant populations, thereby promoting selection pressure towards overall stress resistance. In addition, this nominal concentration would ensure that hexazinone would remain at detectible, phytotoxic levels throughout the experiment despite any degradative or dissipative losses. Enclosures E1 and E3 were assigned as treatments with E2 and E4 as untreated references. The replicates were diagonally opposite each other in the square formed by the four enclosures.
When the enclosure curtains were initially deployed, enclosed water was turbid due to wind-induced mixing of the surrounding channel. Submerged macrophytes were not visible at the time but, as the water column cleared due to reduced winds and stabilization of the bottom by plants, they were observed in all enclosures. By the time of hexazinone treatment, Potamogeton pectinatis and Myriophyllum exalbescens occurred at about the same abundance and degree of patchiness as in the channel. Macrophyte growth continued in untreated enclosures throughout the experiment, to the extent that substrata used for periphyton colonization were barely visible by day 36. Fish removed from the enclosures prior to treatment were mostly young-of-the-year carp (Cyprinus carpio L.) and yellow perch (Perca flavescens Mitchell), brook sticklebacks (Culaea inconstans Kirtland) and fathead minnows (Pimephales promelas Rafinesque). A few fish that escaped capture and fry that developed from egg masses deposited on inner curtain walls by fish trapped by curtain deployment remained alive throughout the experiment in all enclosures.
An effect of enclosure of the Blind Channel was to reduce slightly the turbidity of water due to the windbreak provided by the curtains. The higher water turbidity (Fig. 1A) and planktonic chlorophyll (Fig. 1B) observed in E2 in the first two weeks of enclosure was traced to a curtain breach arising from faulty burial of the lower edge in the sediments. This necessitated its removal and redeployment 24 hours later. Sediment disturbance was minimal during the replacement although some increase in turbulence within the enclosure was unavoidable. To ensure that all enclosures were treated as similarly as possible, all three remaining curtains were also removed and redeployed at the same time. An abrupt, transitory increase in light extinction within the enclosures occurred after curtain redeployment due to mixing with channel water (Fig. 1A). Afterwards, curtain integrity in all enclosures was verified, and differences in light extinction and water chemistry (pH, alkalinity, silicon, ammonia, nitrate, SRP) between enclosures were minor to day 0. Fish removal continued until day 7, by which time no fish of sufficient size to be retained in a minnow trap were caught.
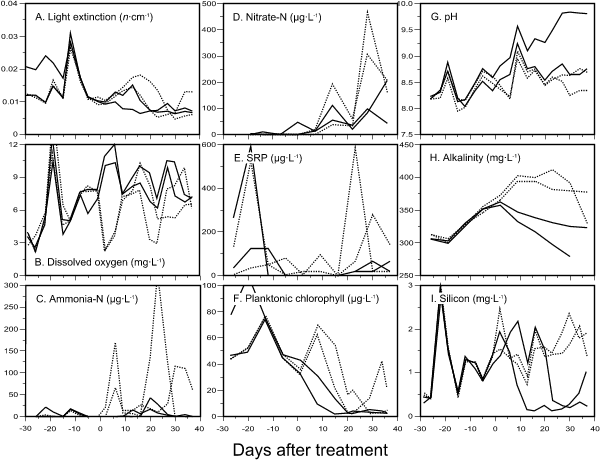
Fig. 1. Changes in physico-chemical parameters in untreated enclosures (solid lines) and hexazinone-treated enclosures (dotted lines) during the experiment. Day 0 = 24 July.
Hexazinone was not detected in the untreated enclosures at any sampling date during the experiment. The initial hexazinone concentration in surface water of E1 was 252 µg·Lą and 231 µg·Lą in E3 (Fig. 2). The fact that these exceeded the predicted nominal concentration may be due to incomplete vertical mixing at the time of first sampling, inaccuracy in estimation of enclosure volume used in calculating the input herbicide volume, or a higher actual hexazinone quantity in the Velpar sample used than was specified on its label (240 g·Lą). Horizontal distribution of the herbicide was assumed to be homogenous since variation between two samples collected 3 m apart from each treated enclosure was usually < 10% of their mean. During the first week after herbicide addition, the absence of thermal profiles in the enclosures (data not shown) suggested that vertical mixing of the water column was complete, thereby distributing hexazinone throughout the enclosure.
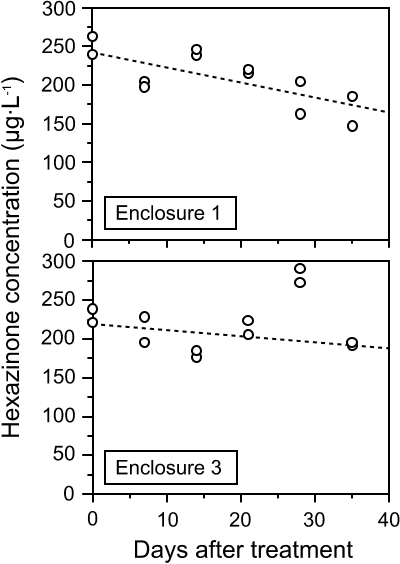
Fig. 2. Changes in hexazinone concentration (µg/L) in treated enclosures E1 and E3 between day 0 and day 35 of the experiment. Points at a given date represent two samples collected from each enclosure. Dashed lines are the zero-order dissipation functions fitted by least-squares linear regression (see text).
As expected from its high water solubility, hexazinone dissipated slowly over time in the treated enclosures. The range of residue concentrations in E1 during the experiment was from 147 to 263 µg·Lą and the dissipation function followed reasonable first-order kinetics (Fig. 2, r = 0.73). An estimated half-life was 60 days. The range of concentration in E3 was from 177 to 291 µg·Lą, but it varied over time more erratically than in E1. The high concentration on day 28 did not correspond to a similar increase in E1, as would be expected if attributeable to a general change at the study site, such as a decrease in water level. An estimated half-life for E3, excluding day 28 data, was 136 days (Fig. 2, r = 0.49).
Hexazinone treatment had a number of immediate and longer-term effects on enclosure water physico-chemistry. Within two days of treatment, the dissolved oxygen (DO) concentration of E1 decreased from 7.8 to 2.4 mg·Lą while E3 decreased from 7.7 to 2.2 mg·Lą (Fig. 1B). The deoxygenation lasted for seven days in both cases; by day 9, levels were only 0.2-0.5 mg·Lą less than untreated enclosures, whose DO levels had increased to > 10 mg·Lą during the corresponding period then decreased. A second period of deoxygenation occurred in E1 between days 20 and 23 and, to a lesser extent, in E3 between days 23 and 30. Variation in DO was closely linked to levels of ammonia (Fig. 1C), nitrate (Fig. 1D) and SRP (Fig. 1E) in treated enclosures. As DO decreased soon after treatment, ammonia increased from below detection to 66 µg·Lą in E1 and 170 µg·Lą in E3. Subsequently, as DO increased, ammonia oxidation lead to increased nitrate which, in turn, decreased as DO decreased and ammonia increased (Fig. 1B,C,D). SRP, which was undetectible in untreated enclosures until the last three weeks of the experiment (Fig. 1E), reaching a maximum concentration of 66 µg·Lą, increased in treated enclosures during periods of low DO, up to 586 µg·Lą, and decreased during periods of higher DO.
Planktonic chlorophyll in the enclosures (Fig. 1F) was directly correlated with light extinction (r = 0.84), with values in E2 higher than in the other three enclosures prior to curtain redeployment. Total planktonic chlorophyll in untreated enclosures decreased from 32-43 µg·Lą at day 0 to < 3 µg·Lą by day 20 and remained low to the end of the experiment. The reoxygenation that occurred in treated enclosures after seven days of treatment coincided with blooms of phytoplankton (62-69 µg·Lą total chlorophyll; Fig. 1F). The blooms, which were dominated by a small ovoid green alga (3-4 µm length) with Nitzschia palea (Kütz.) W.Smith, N. acicularis W.Smith and a few other diatoms as subdominants in both enclosures, increased water turbidity and light extinction (Fig. 1A) as compared to untreated enclosures, where no blooms were noted. The blooms crashed between days 15 and 20, corresponding to the beginning of the second period of deoxygenation and nutrient increase; a recurrence of the bloom in E3 (consisting of the same taxa as before, except that Nitzschia spp. predominated) between days 29 and 34 reversed these trends.
Macrophytes were eliminated from E1 and E3 within seven days of treatment, and no regrowth occurred during the remainder of the experiment.
Persistent effects of herbicide treatment on enclosure pH and alkalinity were observed. The pH of treated enclosures was slightly lower (ca. 0.3 units) than in untreated enclosures (Fig. 1G) while alkalinity was higher (Fig. 1H). Silicon levels in treated enclosures varied between 0.73 and 2.45 mg·Lą with no consistent pattern of change with time, whereas levels in untreated enclosures decreased to < 0.4 mg·Lą (Fig. 1I).
In most cases, changes in water physico-chemistry after day 0 were similar in replicated enclosures. Two exceptions to this occurred in E4, where pH was higher than in other enclosures (Fig. 1G) and silicon decreased earlier than in E2 (Fig. 1I). E3 differed from E1 in that it exhibited two phytoplankton blooms with corresponding changes in ammonia, nitrate and oxygen that occurred once in E1 (Fig. 1).
All substrata became coated with a thin, flocculent brown layer of diatoms within a few days of their placement in all enclosures. The carbon fixation rate (Fig. 3) and chlorophyll concentration (Fig. 4) of periphyton sampled from all enclosures increased exponentially during the first five weeks of substratum colonization (days -27 to -6). Throughout this period, periphyton development on the upper sampled substratum segments (45-50 cm above sediments) was generally higher than on lower segments (15-20 cm above sediments) although diatoms predominated at both positions. In untreated E2 and E4, carbon fixation rate plateaued at about day 0 (four weeks of colonization) although it varied between the week of sampling (Fig. 3) while chlorophyll continued to increase (Fig. 4). Small (ca. 1 mm diameter) gelatinous colonies of Rivularia sp. became abundant on the upper third of each substratum along with diatoms that were more abundant on the lower part. By day 15, substrata had a marly texture and filamentous green algae (Chaeophora sp.) at their top that increased in abundance to day 36. The depth profile seen in early samples were inverted in later samples, with higher carbon fixation rate and chlorophyll observed for the lower substratum segments. Photosynthetic efficiency, the rate of carbon fixation per unit of chlorophyll, was highest for the earliest collected samples (day -27) and, in untreated enclosures, it decreased throughout the experiment. By day 36 in E4, heavy macrophyte cover had resulted in lower periphytic algal biomass on upper portions of substrata amongst the plants; portions below the foliage and those between plant patches supported more biomass and more abundant cyanobacterial colonies and filamentous green algae. Substratum replicate variability was high due to the positions of sampled segments with respect to macrophytes.
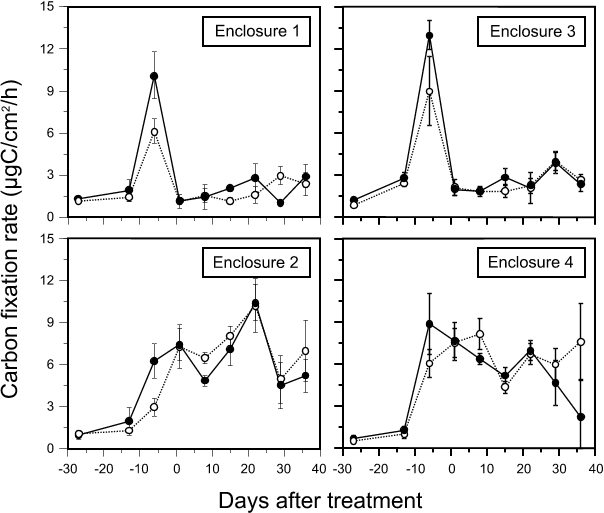
Fig. 3. Changes in periphyton carbon fixation (µgC/cm2/h) on substratum segments collected from untreated enclosures and enclosures treated with 200 µg/L hexazinone (mean ± 1SD, n=6). Solid lines represent segments 45-50 cm above the sediments; dotted lines are for those 15-20 cm above the sediments. Day 0 = 24 July.
Periphyton carbon fixation rate decreased markedly with hexazinone treatment (Fig. 4), to 14% and 19% of levels on day -6 in E1 and E3, respectively. Like in E2 and E4, variation occurred between dates although, in general, carbon fixation gradually increased with time after treatment. The difference in carbon fixation between the two sampling depths was inconsistent between dates, being higher for upper segments only at days 15 and 22. Effects of treatment on periphyton biomass were less abrupt (Fig. 4); a carryover effect of biomass present immediately prior to treatment caused an apparent increase in chlorophyll concentration between day -6 and day 1, which resulted in low photosynthetic efficiency values on day 1 in E1 and E3 (Fig. 5). After this, chlorophyll decreased although the change was not as pronounced as for carbon fixation rates; values at day 8 were 31% and 48% of values on day 1 for E1 and E3, respectively. From day 8, chlorophyll levels in treated enclosures increased slightly over time (Fig. 4). Differences in periphyton chlorophyll with depth after day 8 were small in E1 while in E3, chlorophyll on upper segments was consistently higher than on lower segments (Fig. 7). Photosynthetic efficiency remained constant in both treated enclosures after day 1 and values were lower than for untreated E2 and E4 (Fig. 5). After treatment, globular cyanobacterial colonies that had developed on substrata before herbicide addition remained in place, although they turned from blue/green to brown. Substrata did not have the marliness found on those from untreated enclosures and filamentous green algae were not seen. Substrata remained uniformly brown/black in color to the end of the experiment.
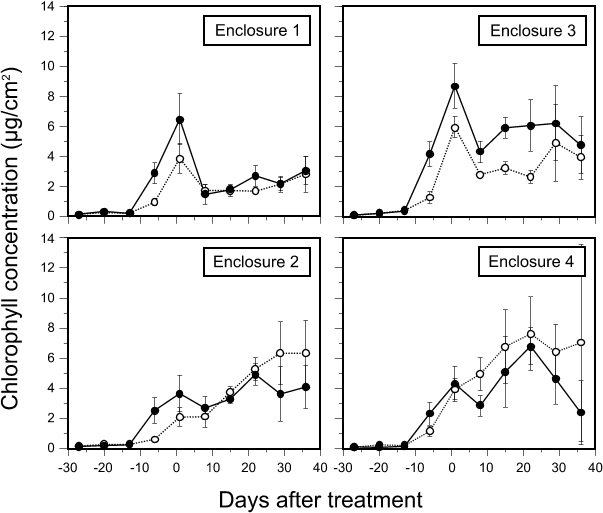
Fig. 4. Changes in periphyton total chlorophyll concentration (µg/cm2) on substratum segments collected from untreated enclosures and enclosures treated with 200 µg/L hexazinone (mean ± 1SD, n=6). Solid lines represent segments 40-55 cm above the sediments; dotted lines are for those 10-25 cm above the sediments. Day 0 = 24 July.
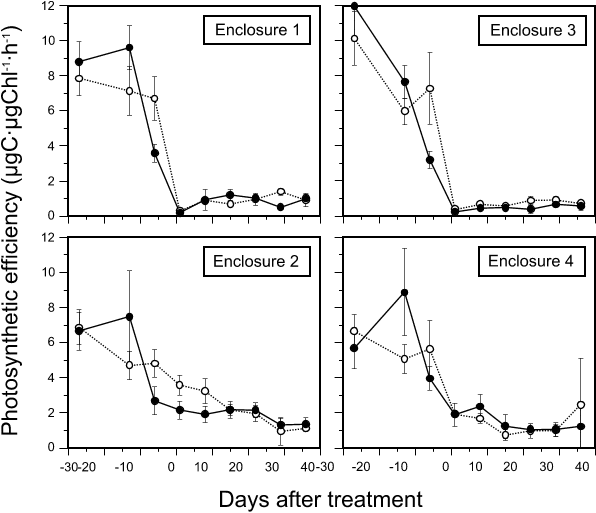
Fig. 5. Changes in periphyton photosynthetic efficiency (µgC/µgChl/h) on substratum segments collected from untreated enclosures and enclosures treated with 200 µg/L hexazinone (mean ± 1SD, n=6). Solid lines represent segments 45-50 cm above the sediments; dotted lines are for those 15-20 cm above the sediments. Day 0 = 24 July.
Evaluation of the significance of changes in periphyton carbon fixation rate and chlorophyll with hexazinone exposure were complicated by the fact that the degree of algal development on substrata prior to treatment was not the same in all enclosures. Summed for all sampling depths and dates before treatment, mean carbon fixation rate in E3 (4.88 µgC·cm˛·hą) was significantly higher (p < 0.01) than in E1 (3.68 µgC·cm˛·hą), which was higher than E4 (3.08 µgC·cm˛·hą) and E2 (2.43 µgC·cm˛·hą). E3 had significantly higher chlorophyll (0.85 µg·cm˛) than the remaining three enclosures, which were not significantly different from each other (0.64, 0.56 and 0.55 µg·cm˛ for E1, E2 and E3, respectively). Consequently, post-treatment differences in absolute levels of carbon fixation and chlorophyll between treated and untreated enclosures were indicative of pre-existing differences as well as to effects of herbicide exposure. Therefore, data were relativized to the maximum observed pre-treatment values of carbon fixation and chlorophyll in each enclosure (which occurred on day -6 on upper segments in all cases) under the rationale that this would show how periphyton development had changed relative to what it had been in each enclosure. Effects of herbicide exposure would be shown by different magnitudes of change in treated enclosures than in untreated enclosures. Considering all post-treatment dates as a whole, carbon fixation was 18% of pre-treatment levels in E1 and 19% in E3 (which were not significantly different from each other at the 1% level), while corresponding values in untreated enclosures were 69% for E4 and 111% for E2. The latter two values were significantly different (p<0.01). For chlorophyll data, however, post-treatment means were 91% of the pre-treatment maximum in E1 and 119% in E3; untreated enclosures were 158% and 225% for E2 and E4, respectively, and all means were significantly different from each other. From these data, it appeared that periphyton carbon fixation was affected by herbicide exposure disproportionately more than chlorophyll concentration.
Several potentially interacting biotic and abiotic factors act as controls on periphyton community development in the Delta Marsh. Intense competition for nutrients with other primary producers, for example, results in levels of major nutrients (e.g., N and P) that are usually below analytical detection limits throughout most of the summer (Goldsborough, unpubl.data). The reducing sediments, which are high in N and P, are the primary source of nutrients to the water column (unpubl.data, Goldsborough and Robinson 1985). Efflux of sedimentary nutrients may be mediated, at least in part, by the rate of their consumption by epipelic algal communities at the sediment/water interface. Macrophytes can utilize nutrients directly from the sediments, but the importance of their subsequent leaching from macrophyte surfaces to associated epiphytic algae is in dispute and may vary with the specific site. Bird feces introduced accidentally to enclosures at the study site have been shown to significantly increase periphyton production (Kotak and Robinson 1991), which clearly demonstrates the limitations imposed on periphyton by nutrient availability. The dominance by heterocystous cyanobacteria (primarily Rivularia sp.) by mid-summer in periphyton at the Delta Marsh, the high nitrogen fixation rates associated with periphyton at this time (Hooper-Reid and Robinson 1978), and the occurrence of the macrophyte Utricularia vulgaris L., which supplements its N budget through carnivory, are also indicators of the low nutrient status in the water column of the marsh. Hann (1991) has demonstrated the importance of herbivory in regulating the structure and overall community biomass of the periphyton community in the Delta Marsh, by selectively grazing erect and pedunculate growth forms in preference to tightly adherent adnate taxa. Water column mixing, which varies seasonally and diurnally with wind activity and the extent of aquatic plant development, also affects periphyton growth by controlling water turbidity which, in turn, determines light availability and nutrient redistribution. Shading and abrasion by dense macrophyte cover also reduces potential periphyton development. Finally, implicit in the increase in structural complexity of periphyton as it develops three-dimensionality over time is the concomitant self-limiting effects on nutrient concentration and light intensity, which are particularly pertinent to those taxa at the bottom of the assemblage.
The differing degree of periphyton development between experimental enclosures prior to hexazinone addition demonstrates the important of these factors. The curtain breach in E2 may have been responsible for lower periphyton biomass and productivity in that enclosure in the first five weeks of substratum colonization (Figs. 3,4). For example, the connection with the channel may have intensified grazing pressure by small fish in E2 by offering them refuge from piscivory. Fish catches in E2 were higher than for other enclosures in the period immediately before curtain redeployment (data not shown). The importance of water turbidity and decreased light availability was seen in all enclosures from the greater periphyton biomass on upper substratum segments than on lower segments when turbidity was high during the first few weeks of the experiment, but which was eliminated or reversed as water clarity increased over time. The increasing architectural complexity of periphyton over time, and its attendant self-limiting effects, was seen by the continual increase in total biomass (chlorophyll) over time as carbon fixation rate remained nearly constant after stabilizing at about day 0. An interpretation for these observations is that measured carbon fixation was due mostly to metabolically active cells near the outside of the mat exposed to higher light and nutrients, while chlorophyll levels reflect the additional component of intact but metabolically less active cells deeper in the mat.
The stabilization of periphyton biomass after an initial period of exponential growth was the net result of competition from increasing macrophyte biomass, possibly increased herbivory from grazers occupying niches among the macrophyte canopy, and self-limitation by the mat itself. The decrease in biomass and productivity of periphyton in E4 towards the end of the experiment suggests that some factor that did not exist to the same magnitude in E2 was controlling periphyton development. While it would be tempting to point to contamination of E4 by hexazinone from either E1 or E3 that bordered it, several lines of evidence suggest this was unlikely. For one, if contamination of E4 had occurred, there is no reason why E2 should not have been similarly affected, yet data from bioassay experiments using periphyton from outside the enclosures incubated in water from the treated enclosures showed no phytotoxicity in E2 over the same period. Given that the decrease in hexazinone concentration over time in treated enclosures was small, and that contamination of the surrounding channel would also have occurred to at least the same magnitude, the amount of herbicide that could have entered E4 would be less than the 10 µg·Lą threshold at which effects on periphyton carbon fixation were observed. One difference between E4 and E2, which may have a bearing on periphyton growth differences between them, is that macrophyte development was more extensive in E4 than in E2. In addition to obvious effects on nutrient availability and shading, the resulting high production in E4 would lead to carbon depletion, consumption of alkalinity and loss of buffering capacity, and increased pH (Fig. 1), which would ultimately limit primary production by periphyton and macrophytes. The earlier depletion of silicon in E4 may be due to growth of epiphytic diatoms which were abundant at the study site. The periphyton in E4 was, by the end of the experiment, predominated by filamentous green algae whereas diatoms were more abundant in E2. The difference in periphyton development in the two untreated enclosures by day 36 would lead one to question the replicability of enclosures that, although intended by their larger size to minimize effects of enclosure on water chemistry, also tended to promote heterogeneity by, for example, encompassing macrophyte patchiness that is normal in the Blind Channel. I contend that despite these concerns, such effects are a necessary tradeoff for greater realism in assessing the potential effects of herbicide contamination of the marsh as a whole, where heterogeneity is inherent. The single concession to enclosure uniformity was to remove as many fish as possible since it would have been difficult, if not impossible, to ensure that their density was the same in all enclosures. Any attempt to manipulate fish density would have added one more variable to an already complex system.
Effects of hexazinone on enclosure water chemistry were similar to those from previous studies using the related triazine herbicides atrazine, simazine and terbutryn in enclosures (Goldsborough and Robinson 1983, Hamilton et al. 1989). It is likely that deoxygenation was due to decreased photosynthesis and increased respiration as algal and macrophytic biomass decayed. However, the decrease in oxygen was shorter and less severe than in previous experiments with simazine at the study site, where complete anaerobiosis persisted for as long as 22 days (Goldsborough and Robinson 1985). The increase in N and P with herbicide treatment can be attributed, in part, to their release and mineralization during plant decomposition. Enhanced sediment efflux under anoxic conditions due to reduction in epipelic assimilation with herbicide phytotoxicity may also be important (Jansson 1980, Goldsborough and Robinson 1985). Increases in dissolved inorganic carbon in triazine-treated enclosures, relative to untreated enclosures, have been reported by others (e.g., Herman et al. 1986, Hamilton et al. 1989) reflecting decreases in carbon assimilation with photosynthetic inhibition.
Hexazinone acted as an additional chemical stressor on the periphyton community, reducing both its biomass and productivity within seven days of exposure. The response of these two parameters differed, however, with effects on carbon fixation rate more immediate and of greater magnitude than those on chlorophyll concentration. Two explanations may be offered for the discrepancy. One is that carbon fixation rate, as a metabolic indicator, responded to a photosynthetic inhibitor more quickly than biomass, since measurements of chlorophyll do not differentiate metabolically active components of the community from those that are inactive or dead. This may explain the residual effects on chlorophyll seen at day 1 (Fig. 4). Alternatively, carbon fixation rate may not be the only means of production employed by periphyton experiencing herbicide-induced stress; they may resort to facultative heterotrophy with the result that total biomass (as indicated by chlorophyll, assuming that myxotrophs retain it) may be more representative of community function that total inorganic carbon fixation. Since it is not possible to determine which of these possibilities is more likely for the present data, one is forced to the conclusion that each parameter offers unique information in evaluating periphyton response to stress.
It is expected that the degree of tolerance to a given stressor by members of a natural algal assemblage follows a normal curve ranging from extreme stress-sensitivity to extreme stress-tolerance, with most taxa being intermediate (Cairns and Niederlehner 1989). It is not unexpected, therefore, that carbon fixation should continue at high hexazinone concentrations under laboratory exposure of that carbon fixation of natural periphyton should not decline to zero except at extremely high levels of the stressor. Goldsborough and Robinson (1988) showed that the EC50 concentration for the effects of simazine on carbon fixation increased for periphyton from a simazine-treated enclosure. Although both carbon fixation and chlorophyll in E1 and E3 did increase between days 8 and 36, this is consistent with continued growth by a smaller, stress-tolerant subset of the stressed community. The fact that the rate of community growth with time was much less than that seen in all enclosures prior to introduction of the herbicide argues that any increase in community resistance was insufficient to allow the community to continue growth at rates approaching those in the absence of herbicide stress. Therefore, the data do no support a hypothesis that periphyton resistance would increase with continued exposure to the stressor.
Phytoplankton has usually been considered to be a comparatively smaller contributor to total primary production in channels of the Delta Marsh than macrophytes and periphyton, whose growth is profuse (e.g., Hooper and Robinson 1976). High water turbidity is generally attributed to suspended sediments, and when the bottom becomes stabilized later in the summer, water clarity is high. Water samples collected in the marsh invariably contain tychoplanktonic taxa arising from suspended epipelon and epiphyton. These cells, along with suspended detritus, may contribute to high chlorophyll levels in the water column (35-80 µg·Lą in 1991; Goldsborough, unpublished data). Therefore, it is of practical significance that phytoplankton flourished in herbicide-treated enclosures whereas macrophytes, and to some extent, periphyton did not. Other investigations of the response of phytoplankton to triazine herbicide exposure have noted increases in the abundance of selected taxa, which has been used as evidence for development of functional (i.e. photosynthetic) resistance. DeNoyelles et al. (1982) reported increases in the density of Mallomonas sp. and Cryptomonas spp. in ponds treated with 500 µg·Lą atrazine. Similar proliferation of cryptomonads in response to 100 µg·Lą atrazine has been reported by Hamilton et al. (1989). A possible explanation for these observations that has apparently been overlooked is that cryptomonads, which are known to be facultatively phagotrophy (e.g., Tranvik et al. 1989), may be responding via a nutritional mode shift to increased bacterial abundance with increases in DOC arising from decaying autotrophs killed by herbicide exposure. By contrast, although detailed community structural analyses of phytoplankton in these enclosures are unavailable, circumstantial evidence, including the predominance of taxa with unknown heterotrophic potential and with documented affinities for high inorganic nutrient conditions (e.g., Nitzschia spp.) and the apparent correlation between reoxygenation, depletion of inorganic nutrients in the water column and phytoplankton chlorophyll, argues that these phytoplankton were autotrophic and had greater physiological tolerance to hexazinone than other algae. The blooming of herbicide-tolerant phytoplankton in both treated enclosures (and their resurgence in E3), in the absence of significantly increased resistance in periphytic algal communities begs the question as to why resistance would increase in phytoplankton, allowing them to become abundant, and not in periphyton. It may be that phytoplankton can respond more quickly and more abundantly to nutrient enrichment resulting from herbicide exposure. They may also occupy a larger niche than periphyton because they inhabit a water column that is generally well mixed and chemical homogenous and they are distributed by water movement. Periphyton, by contrast, are necessarily restricted by substratum availability (which, in treated enclosures, consisted entirely of artificial substrata, the enclosure walls, and the sediments), by the abundance of niches existing in a three-dimensional matrix defined by physical, chemical and biological gradients, and by the fact that most periphytic taxa are weakly motile or nonmotile and might disperse slowly to newly available substratum. In addition, once developed, the phytoplankton bloom may further stress the periphyton community through shading, and by decreasing nutrient supplies through competitive assimilation.
If it is assumed that members of the Delta Marsh ecosystem have evolved to maximize their ability to utilize available energy resources, the shift in the composition of primary producers may impact on the availability and suitability of food resources to grazers and the higher order consumers that depend on them. There have been several documented cases where anthropogenic factors have altered the composition of aquatic primary producers with consequent impacts on herbivory and energy availability to consumers. Two examples include the progressive elimination of macrophytes and their replacement by phytoplankton in lakes undergoing cultural eutrophication (e.g., Phillips et al. 1978), and the proliferation of metaphytic green algal mats in experimentally acidified lakes (Howell et al. 1991). If, in either case, the favoured primary producers were less easily grazed than the algae they replaced, the entire ecosystem would be affected negatively.
Future work will attempt to evaluate the generality of the conclusions reached here, that phytoplankton may replace periphyton and macrophytes at sublethal levels of herbicide stress, that effects of stress on chlorophyll and carbon fixation differ, that increases in periphyton resistance are minor and transitory, and that stress effects, at least in the context of a sublethal exposure of < 6 weeks, are reversible. Moreover, since it is rare, if ever, that a single environmental factor is responsible for structuring living communities, observed periphyton productivity and biomass is likely the net response to multiple simultaneously interacting stressors and stimulants. Before these data may contribute to a generalized model of the response by aquatic primary producers to herbicide stressors, data on the importance of these potential interactions will be needed.
Financial support for this research was provided by an Operating Grant from the Natural Sciences and Engineering Research Council of Canada. I gratefully acknowledge the field assistance of Mike Forster and Maria Zbigniewicz, and the researchers and staff of the University Field Station (Delta Marsh). D. Rourke assisted with enclosure construction. DuPont Canada Inc. kindly provided Velpar L(r) used in this study. N. Bertrand, R. Greenhalgh and the staff of Xenos Laboratories Inc. performed residue analyses and Simplot Canada Ltd. performed nitrate analyses.
Anderson, L.W.J. 1981. Control of aquatic weeds with hexazinone. J. Aquat. Pl. Mgmt 19: 9-14.
American Public Health Association (APHA). 1980. Standard methods for the examination of water and wastewater. 15th ed. Washington, DC. 1134 p.
Blanck, H. and S-Ĺ. Wängberg. 1988. Induced community tolerance in marine periphyton established under arsenate stress. Can. J. Fish. Aquat. Sci. 45: 1816-1819.
Cairns, J.,Jr. and B.R. Niederlehner. 1989. Adaptation and resistance of ecosystems to stress: a major knowledge gap in understanding anthropogenic perturbations. Specul. Sci. Technol. 12: 23-30.
Campbell, R.A. 1990. Herbicide use for forest management in Canada: where we are and where we are going. Forestry Chronicle 8: 355-360.
deNoyelles, F., W.D. Kettle, and D.E. Sinn. 1982. The responses of plankton communities in experimental ponds to atrazine, the most heavily used pesticide in the United States. Ecology 63: 1285-1293.
Goldsborough, L.G. and M.J. Forster. Effect of exposure period and architectural development on the toxicity of a herbicide to a periphyton community. Submitted to Aquatic Toxicology, July 1992.
Goldsborough, L.G. and G.G.C. Robinson. 1983. The effect of two triazine herbicides on the productivity of freshwater marsh periphyton. Aquat. Toxicol. 4: 95-112.
Goldsborough, L.G. and G.G.C. Robinson. 1985. Effect of an aquatic herbicide on sediment nutrient efflux in a freshwater marsh. Hydrobiologia 122: 121-128.
Goldsborough, L.G. and G.G.C. Robinson. 1988. Functional responses of freshwater periphyton to short simazine exposures. Verh. Internat. Verein. Limnol. 23:1586-1593.
Hamilton, P.B., D.R.S. Lean, G.S. Jackson, N.K. Kaushik and K.R. Solomon. 1989. The effect of two applications of atrazine on the water quality of freshwater enclosures. Env.Poll. 60:291-304.
Hann, B.J. 1991. Invertebrate grazer-periphyton interactions in a eutrophic marsh pond. Freshw. Biol. 26: 87-96.
Herman, D., N.K. Kaushik, and K.R. Solomon. 1986. Impact of atrazine on periphyton in freshwater enclosures and some ecological consequences. Can. J. Fish. Aquat. Sci. 43: 1917-1925.
Hooper, N.M. and G.G.C. Robinson. 1976. Primary production of epiphytic algae in a marsh pond. Can. J. Bot. 54: 2810-2815.
Hooper-Reid, N.M. and G.G.C. Robinson. 1978. Seasonal dynamics of epiphytic algal growth in a marsh pond: composition, metabolism, and nutrient availability. Can. J. Bot. 56: 2441-2448.
Howell, E.T., M.A. Turner, R.L. France, M.B. Jackson and P.M. Stokes. 1991. Comparison of Zygnematacean (Chlorophytal) algae in the metaphyton of two acidic lakes. Can. J. Fish. Aquat. Sci. 47: 1085-1092.
Jansson, M. 1980. Role of benthic algae in transport of nitrogen from sediment to lake water in a shallow clearwater lake. Arch. Hydrobiol. 89: 101-109.
Kotak, B.G. and G.G.C. Robinson. 1991. Artificially-induced water turbulence and the physical and biological features within small enclosures. Arch. Hydrobiol. 122: 335-349.
Marker, A.F.H., C.A. Crowther, and
R.J.M. Gunn. 1980. Methanol and acetone for estimating
chlorophyll a and phaeopigments by spectrophotometry.
Arch. Hydrobiol.
Beih. 14: 52-69.
McQueen, D.J., J.R. Post, and E.L. Mills. 1986 Trophic relationships in freshwater pelagic ecosystems. Can. J. Fish. Aquat. Sci. 43: 1571-1581.
Muir, D.C.G. and N.P. Grift. 1987. Herbicide levels in river draining two prairie agricultural watersheds. J. Env. Sci. Hlth B22: 259-284.
Phillips, G.L., D. Eminson and B. Moss. 1978. A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters. Aquat. Bot. 4: 103-126.
Power, M.E. 1992. Top-down and bottom-up forces in food webs: do plants have primacy? Ecology 73: 733-746.
Reupert, R., E. Ploger and G. Brausen. 1990. HPLC determination of 29 controlled herbicides in water supplies. Hewlett-Packard Application Note.
SAS Institute Inc. 1986. SAS / Stat Guide for Personal Computers, Version 6 Edition. SAS Institute, Cary, NC, 378 p.
Stainton, M.P., M.J. Capel and F.A.J. Armstrong. 1977. The chemical analyses of fresh water. 2nd ed. Can. Fish. Mar. Serv. Misc. Publ. 25: 180 p.
Tranvik, L.J., K.G. Porter, and J.M. Sieburth. 1989. Occurrence of bacterivory in Cryptomonas, a common freshwater phytoplankter. Oecologia 78: 473-476.
Weed Science Society of America (WSSA). 1983. Herbicide handbook. 5th ed. Weed Science Society of America. Champaign, Illinois. 515 p.
Williamson, D.A. 1988. Hexazinone residues in surface and groundwater at two sites within Agassiz Provincial Forest, Manitoba, Canada. Wat. Poll. Res. J. Can. 23: 434-449.